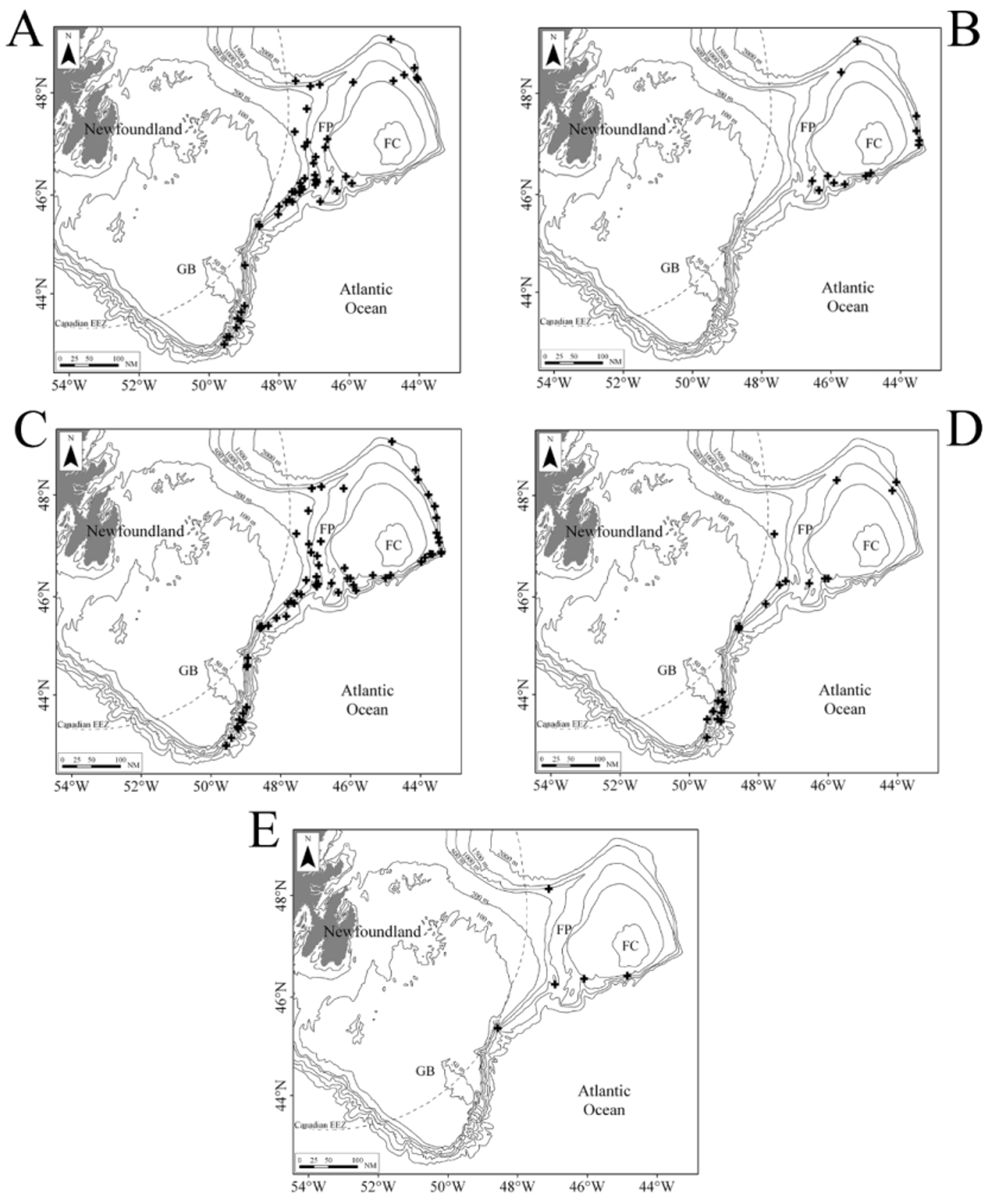
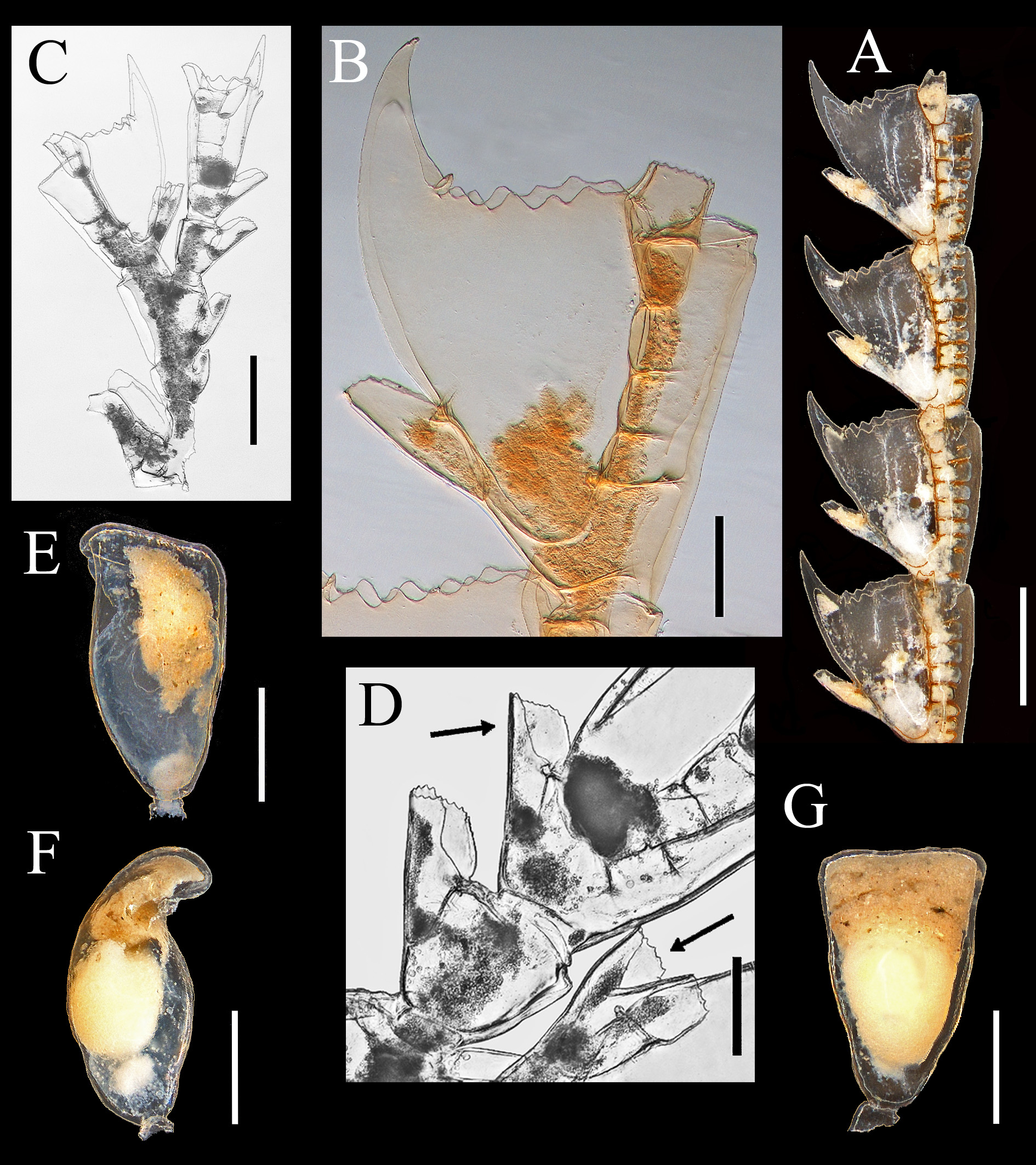
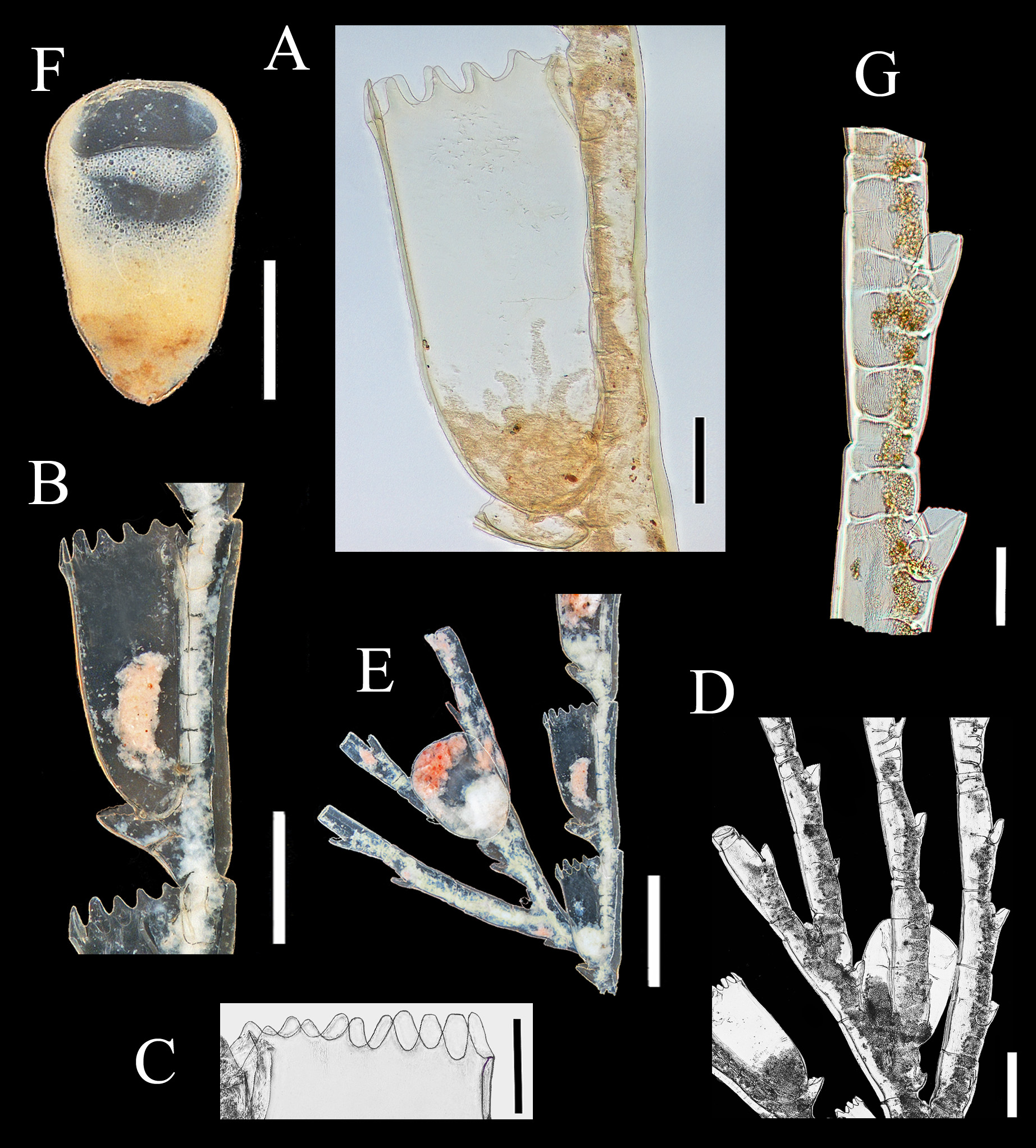
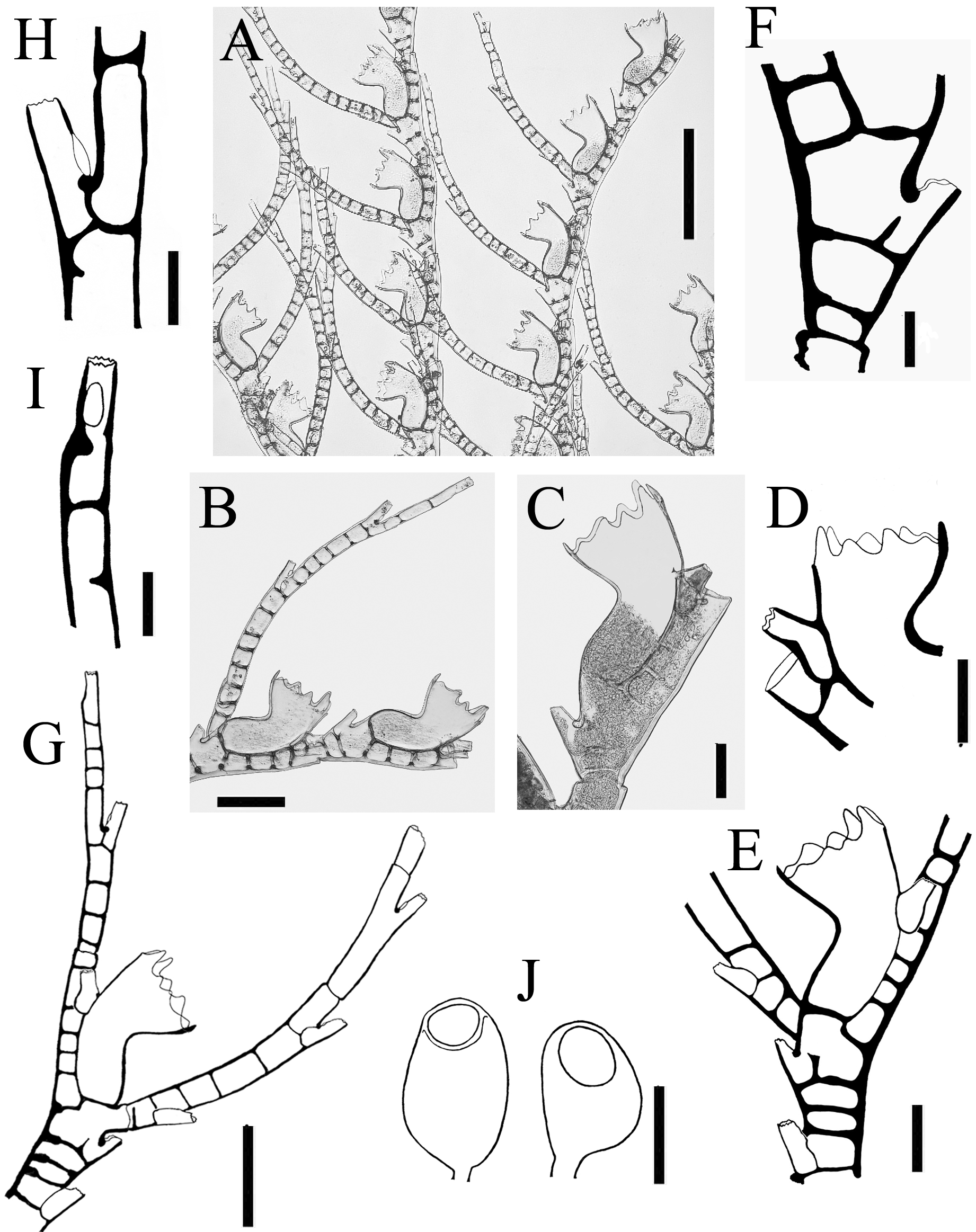
Discussion The discovery of five aglaopheniid species off the east coast of Newfoundland, two of them not recorded previously in the western North Atlantic, is noteworthy. Aglaopheniids are predominantly a warm-water group of hydroids (Calder 1997 a), and their diversity is low in cold Atlantic waters (Fraser 1944). Only eight species are known previously in the region from 40 ºN (New York Bight) northwards to the Arctic Ocean in northeastern North America (Table 1), and most of those have been reported infrequently. However, biodiversity of the deep-water fauna in this region is comparatively less well known than at equivalent latitudes in the northeast Atlantic. For example, 12 species of aglaopheniids (11 of Cladocarpus Allman, 1874 and one of Lytocarpia Kirchenpauer, 1872), were reported by Schuchert (2001) from waters around Iceland, and a greater number are known from 40 ºN northwards in European waters (Cornelius 1995; Altuna 2007). Based on current knowledge, species richness in the family therefore appears to be higher at a given latitude in the northeastern than in the northwestern Atlantic. The disparity in species richness, particularly at shallow depths, may be due in part to differences in sea temperatures at a given latitude in North America and Europe. However, there has also been unequal sampling effort and taxonomic work in the two regions. For instance, Kramp (1943, table 2), listed six species from the west coast of Greenland, while only three were known previously from eastern Canada. Aglaophenopsis cornuta, Cladocarpus formosus, and Cladocarpus integer are quite widely distributed (although infrequently reported) from the Davis Strait and Baffin Bay regions southwards at least to Newfoundland. In contrast, Cladocarpus diana and Nematocarpus ramuliferus are new records for the northwestern North Atlantic. Both occur on the Flemish Cap, a location where all species dealt with in this paper were recorded (Table 11). The Flemish Cap is a plateau of approximately 200 km width, with depths of less than 150 m at its center. Situated eastward of the Grand Banks of Newfoundland, it is separated from those banks by the approximately 1200 m deep Flemish Pass (Figure 1). The Flemish Cap is located within an area of transition between cold subpolar waters and is influenced by fluctuations in the Labrador Current and in the North Atlantic Current (Gil et al. 2004). Compared with the Grand Banks, the Flemish Cap supports higher primary and secondary productivity due to its hydrodynamic conditions (Maillet et al. 2005). The mix of different currents in the area (Gil et al. 2004), a highly oxygenated environment rich in nutrients (Stein 2007), and greater substratum heterogeneity on the Flemish Cap compared with adjacent areas may explain differences in its faunal composition. It is a poorly studied area, and one in which rare and seldom-recorded species of other animal groups, such as deepsea corals (Murillo et al. 2011), are not uncommon. According to those authors, species exist on the Flemish Cap that are not usually recorded in deep-waters off the Canadian coast, and species richness is higher compared with more extensive areas immediately to the west. Such faunal differences probably also exist in the hydroid fauna, but no investigations have been undertaken to support this assumption. As noted in the Introduction, genera of Aglaopheniidae are differentiated mainly on the basis of gonosomal characters. Of eight genera recognized in the family worldwide by Bouillon et al. (2006), Cladocarpus is second in terms of diversity, with 66 species of mostly deep-water forms. Two approaches have been adopted in the taxonomy of this group of aglaopheniids. One of these, adopted by Bouillon et al. (2006) and others, is to recognize a broadly defined genus Cladocarpus, treating it essentially as a collective group until questions of relationships become clearer. The second approach is to refer Cladocarpus - like hydroids to several genera, distinguishing them largely by differences in morphology of their phylactocarps that are thought to reflect relationships. Following this approach, Ramil & Vervoort (1992) distinguished between species having a phylactocarp homologous to the axis (Cladocarpus s. l.) from those in which the rachis is similar structurally to the hydrocladia (Streptocaulus). Their proposals have not been widely accepted because of the existence of intermediate material with respect to this character (see Schuchert 2001). Given the present limited knowledge, actual relationships thus remain obscure. Phylogeny of the group should become clearer as cladistic studies and molecular analyses are undertaken. Particular attention in such investigations must be given to the type species of the various putative genera. Whichever taxonomic approach is followed now, revisions in classification and nomenclature can be expected, although we maintain that fewer changes will be likely to take place when the second alternative is adopted. We therefore support recognizing various genera within this group based on available evidence, rather than simply assigning a large and diverse assemblage of species to an excessively broad Cladocarpus. The genus is already species-rich, and adding many dissimilar taxa to it overextends its scope and renders it more difficult to conceptualize and refine. In our opinion, several nominal genera included in the synonymy of Cladocarpus sensu lato will almost certainly be recovered as valid based on differences in structure of the phylactocarp. In any case, reproductive structures in species of Cladocarpus, as conceived by Bouillon et al. (2006), are heterogeneous. All five species recorded herein were assigned to the same genus (Cladocarpus) in that work, notwithstanding significant variation in morphology of their gonosomes (and in one case colony structure). We conclude that the five species are referable to three genera, viz. Cladocarpus, Aglaophenopsis, and Nematocarpus. The taxonomy of Cladocarpus formosus is unmistakable, as it is the type species of the genus. Cladocarpus diana is also referable to Cladocarpus based on characters of its reproductive structures. The generic identity of Cladocarpus integer is less clear. Its phylactocarp is an unbranched structure and as such is dissimilar to that of C. formosus. Still, species with both branched and unbranched phylactocarps are currently assigned to Cladocarpus, and we include C. integer in that genus for the time being. The status of Nematocarpus, and the generic affinities of its sole species N. ramuliferus, are unclear. Nematocarpus is closely related to Aglaophenopsis, and the two were regarded as congeneric by Calder (1997 a). Hydrothecae and ramuli / phylactocarps of Aglaophenopsis hirsuta Fewkes, 1881 (type species of Aglaophenopsis) and Nematocarpus ramuliferus (Allman, 1874) (type species of Nematocarpus) are quite similar, as apparent in illustrations of A. hirsuta by Vervoort (1972: 206) and N. ramuliferus by Broch (1918: 76) and Schuchert (2001: 144). Gonothecae and overall colony forms of the two are also similar. However, ramuli of N. ramuliferus are not restricted to the proximalmost cormidia, as occurs with the phylactocarps of Aglaophenopsis and Cladocarpus, and the gonothecae are borne only on the apophyses of the main tube of stem and branches, and not on the ramuli. Hence, ramuli do not appear to function primarily as reproductive structures protecting the gonothecae, as is the case with phylactocarps of A. hirsuta. Hydroids belonging to the genus Calvinia (Halopterididae) are recognized as distinct based on possession of such accessory tubules, and the same character provides justification for recognition of Nematocarpus. Thus, Nematocarpus is upheld here as a valid genus, containing its type species, N. ramuliferus. Some seven species have been assigned to Aglaophenopsis, with six of them occurring in the North Atlantic (Table 10). The phylactocarp in the type species, A. hirsuta Fewkes, 1881, arises in place of the mesial nematotheca (Fewkes 1881). Vervoort (1972) noted that it was borne on one of the funnels when the nematotheca is bifid. This difference was clearly stated by Bale (1886), who noted that “ the phylactocarps of Aglaophenopsis are modified from the mesial nematophore of the proximal hydrothecae, while in Cladocarpus they are independent structures, and the genus must rest on this distinction ”. The emended diagnosis of the genus by Fraser (1944) asserts that the gonosome is “ protected by phylactogonia that appear to be modified mesial nematophores ”. However, Bogle (1975) did not mention phylactocarps as modified mesial nematothecae in her diagnosis of the genus, stating: “ phylactogonia arising singly from the proximal internode of unmodified hydrocladia from an area to the side and slightly above the mesial nematotheca ”. In fact, not all species referred to Aglaophenopsis have modified mesial nematothecae (see Table 10). Bogle emphasized the presence of hydrothecae on the phylactocarps as a distinction from Cladocarpus in her key to genera, and proposed other characters for differentiation. Accordingly, we are uncertain about the correct generic assignment of Cladocarpus cornutus Verrill, 1879 (and Aglaophenopsis bonnevieae, which we regard as congeneric with A. cornuta). While both species are usually referred to Aglaophenopsis, we question whether they are congeneric with A. hirsuta, type species of that genus, because their phylactocarps are different. In both A. cornuta and A. bonnevieae they are forked (‘ Y-shaped’, or even alternately branched in large colonies of A. bonnevieae) and consistently hydrothecate, usually throughout (e. g., see Kramp 1932 a: 59; Schuchert 2001: 138) (Figure 2 C). Instead, those of A. hirsuta (see Nutting 1900: pl. 29, fig. 12; Vervoort 1972: 206) are unbranched, and hydrothecae are of sporadic occurrence. Gonothecae also seem to differ, especially those of A. cornuta, apertures of which are lateral and oval in shape, rather than terminal and circular as in A. hirsuta. These two species (A. cornuta and A. bonnevieae) do not correspond readily to any of the described genera in the tribe Cladocarpini Calder, 1997 a, and a case could be made for a new genus to accommodate these two. For now, however, we have taken a conservative approach and have retained them in Aglaophenopsis. In Cladocarpus formosus, type species of the genus, the mesial nematotheca on thecate internodes is almost entirely adnate to the abcaulinar wall of the hydrotheca except on the first internode, where there is a distinct gap between distal end of nematotheca and hydrothecal base (Figure 4 F). Phylactocarps arise from this area. The same morphology is seen in Cladocarpus integer (Figure 5 E, F) and Cladocarpus diana, with the mesial nematotheca being in a different position on the first thecate internode than on the others. In C. diana, however, the gap is not so apparent, with the phylactocarp arising clearly from the lateral side (Figure 3 E). In Aglaophenopsis cornuta, the mesial nematotheca of the first cormidium is adnate to the hydrotheca, but it is displaced to the side as seen in frontal view. In all four of these species, the position of the mesial nematotheca with respect to the hydrotheca is different on the proximalmost internode of the hydrocladium than on the rest. In Nematocarpus ramuliferus, however, there is no such difference. Phylactocarps occur only on the first thecate internode in Cladocarpus s. l., and a gap exists between the mesial nematotheca and hydrotheca on this internode, even in species having nematothecae that are adnate to hydrothecae on other hydrocladial internodes. This difference also occurs in sterile colonies. Gonothecae in A. cornuta and C. formosus occur on both apophyses (1 – 2 gonothecae) and on the phylactocarp. In C. diana, they arise from the phylactocarp. Gonothecae in hydroids of C. integer examined here were borne on the phylactocarp, although colonies with gonothecae on the stem, but lacking phylactocarps, are known in the species (Broch 1918). In N. ramuliferus, gonothecae occur only on apophyses of the main tube. Notably, colonies of A. cornuta and C. formosus with gonothecae, but with no phylactocarps, were observed in our material. Such colonies have also been reported in Cladocarpus campanulatus Ritchie, 1912 (Schuchert 2001). This suggests that the absence of phylactocarps in fertile material is probably much more common within the Cladocarpini than previously thought. The most abundant species in our collections was C. formosus. Cladocarpus integer was found at the shallowest depth (119 m), while the deepest recorded species were A. cornuta and C. formosus (1885 m). The latter two species also had the widest bathymetric range (1645 m). Hydroids of exceptionally large size were collected in some of the species (Table 9). The largest colony, a specimen of C. integer, was 78.0 cm high. The largest ones of C. formosus and A. cornuta were 53.5 cm and 28.7 cm high, respectively. Cladocarpus diana and N. ramuliferus are small hydroids not exceeding 7.0 cm in height. Of particular research interest, the habitat provided by several deep-water anthozoans has been emphasized only recently. The question arises whether large hydroid colonies, such as those found here, might also provide important substrate and shelter. However, sessile epibionts and vagile invertebrates were scarce amongst the colonies examined. Observed associates included other hydroids, alcyoniids, brachiopods, barnacles, bryozoans, and caprellid amphipods. Such a limited number of associates suggests that these hydroids may have effective antifouling defenses. Identification key to aglaopheniid hydroids reported from the western North Atlantic between 40 ºN and Baffin Bay, including western Greenland
Altuna, Álvaro, Murillo, Francisco J., Calder, Dale R. (2013): Aglaopheniid hydroids (Cnidaria: Hydrozoa: Aglaopheniidae) from bathyal waters of the Flemish Cap, Flemish Pass, and Grand Banks of Newfoundland (NW Atlantic). Zootaxa 3737 (5): 501-537, DOI: 10.11646/zootaxa.3737.5.1