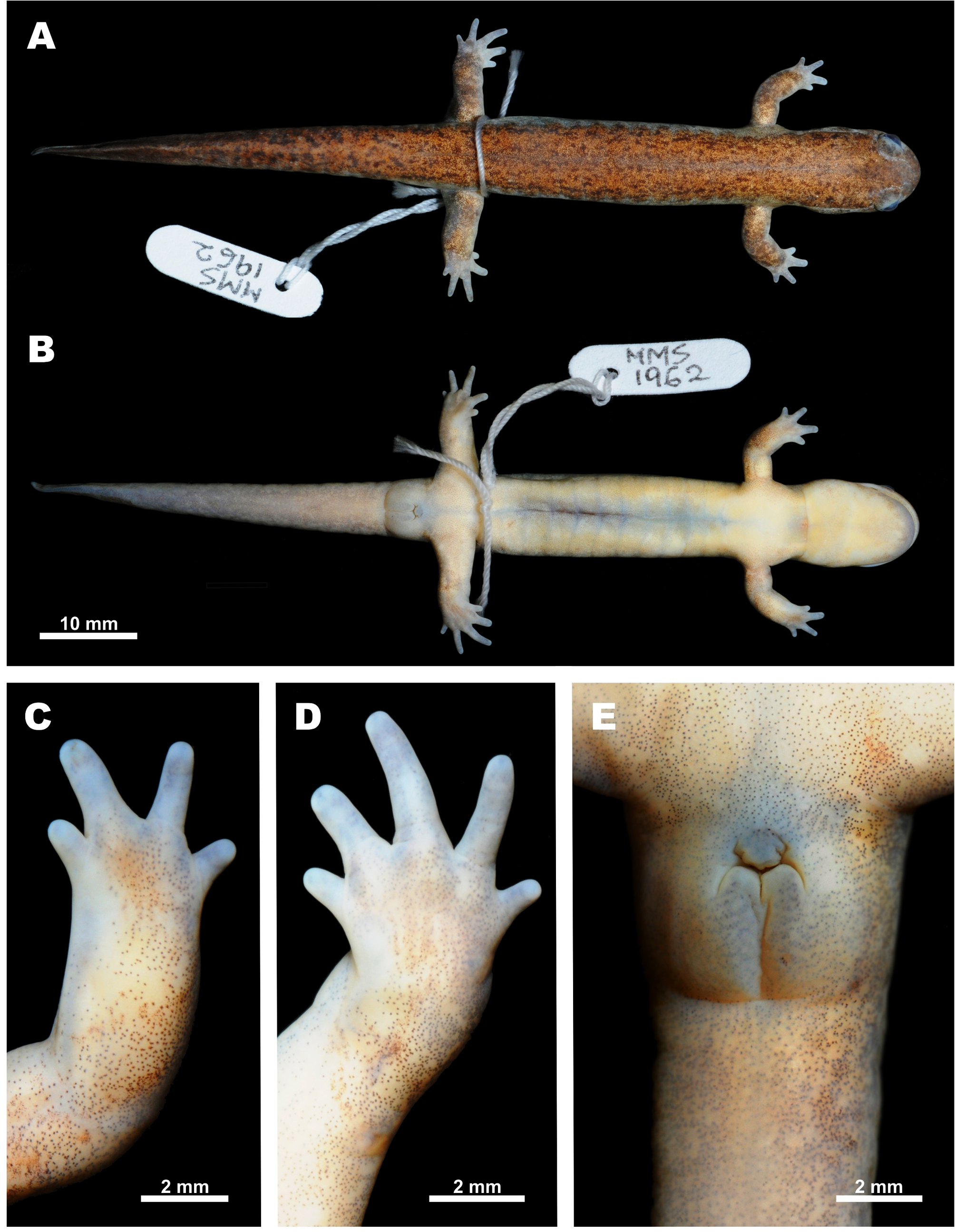
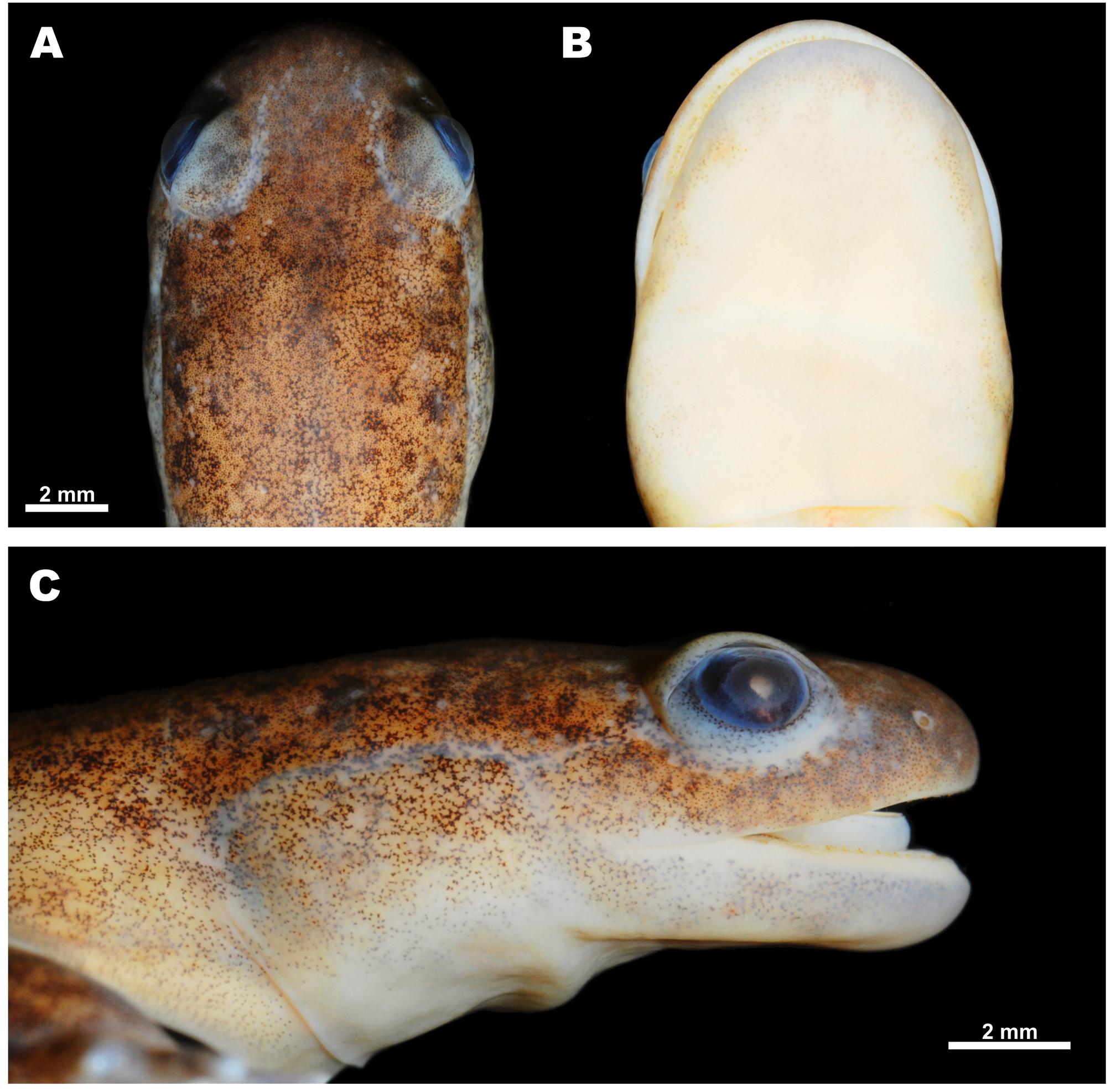
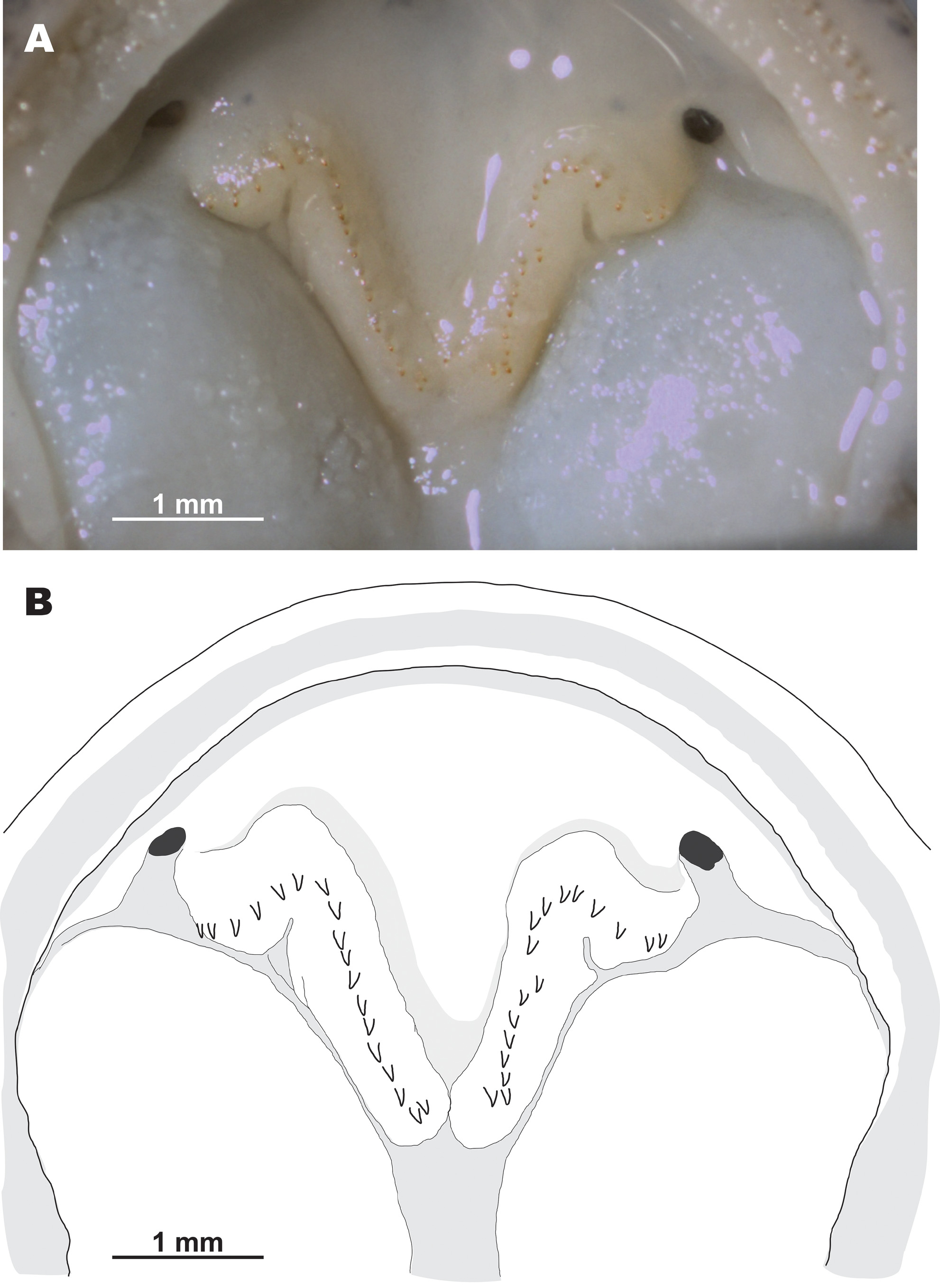
Distribution, habitats and natural history. To date, the new species has been found in four localities within the southeastern part of Jeollanam-do, South Korea: Goheung, Suncheon, Bosoeung and Yeosu areas (Fig. 9, localities 1 – 6). Specimens from these localities were used in the genetic study of Baek et al. (2011 a, 2011 b) and their taxonomic assignation to Hynobius unisacculus sp. nov. was confirmed by mtDNA sequence analysis. However, Chang et al. (2011) reported several other populations of the H. leechii species complex on adjacent offshore islands along the southeastern coast of Jeollanam-do (Fig. 9, localities 7 – 12); the taxonomic status of these populations requires further study. The new species reproduces in still waters of ditches or rice paddies, after the breeding season, adults move onto surrounding land to hide in forest litter or other shelters (see Fig. 7). According to the data of Chang et al. (2011), on Oenaro-do Island, Hynobius unisacculus sp. nov. is sympatric with Bufo gargarizans Cantor, Hyla japonica Günther and Pelophylax nigromaculatus (Hallowell). On Gogeum-do Island, what is assumed to be the new species (identified as H. leechii) was recorded in sympatry with the above mentioned anuran species, and also with Bombina orientalis (Boulenger), Rana coreana (Okada), and the introduced Lithobates catesbeianus (Shaw). On Geomun-do Islands (Dong-do and Seo-do) the population tentatively assigned to the new species represents the only amphibian species recorded (Chang et al. 2011). Comparisons. Among the 35 currently recognized species of the genus Hynobius (see Frost 2016), Hynobius turkestanicus Nikolskii appears to be an enigmatic taxon (see Kuzmin 1999) and is likely not a member of this genus. The rest of the species are grouped into three discrete groups, differing in morphology, phylogenetic position, chromosome structure, breeding ecology, and natural history. The stream-breeding or lotic Hynobius found in Taiwan and Japan, as well as H. (Satobius) retardatus, found in Hokkaido, are markedly distinct from the lentic-breeding species of Hynobius, reproducing mostly in still waters and inhabiting eastern and northeastern China, the Korean Peninsula and Japan. The new species, H. unisacculus sp. nov., is both most morphologically and most molecularly similar to other lentic-breeding species of Hynobius from the Korean Peninsula: H. leechii, H. quelpaertensis and H. yangi, and comparisons with these taxa and other lentic-breeding Hynobius species of the region appear to be the most pertinent. Morphological comparison of H. unisacculus sp. nov. with three other species of Korean Hynobius are given in Tables 2 and 3. The new species is a small-sized Hynobius (up to 61 mm SVL; mean SVL 49.0 mm in males and 51.4 mm in females), with short limbs and slender body. In SVL, post hoc analyses of one-way ANOVA revealed that H. unisacculus sp. nov. was not significantly different from H. yangi from Busan, but was significantly much smaller than H. quelpaertensis from Jeju Island and H. leechii from mainland Korea (one-way ANOVA, df = 3, F = 42.49, P <0.05; Duncan test, P <0.05). The new species can be further differentiated from H. quelpaertensis by its much shorter tail with mean TL / SVL ratio for males being 0.79 ± 0.01 and for females 0.73 ± 0.02 (versus 0.90 ± 0.02 in males and 0.79 ± 0.02 in females of H. quelpaertensis); these differences are statistically significant (TL, one-way ANOVA, df = 3, F = 9.360, P <0.01). H. unisacculus sp. nov. has relatively shorter limbs and longer body than H. quelpaertensis: between adpressed limbs in the new species there is always a wide gap of - 2.4 costal folds in males and - 2.5 costal folds in females (ranging from - 3.5 – - 1.5), whereas in H. quelpaertensis the adpressed limbs often touch or overlap and the CGBL value is 0 in males and - 1.3 in females (ranging from - 1.5 – 1.0). H. unisacculus sp. nov. has a lower number of vomerine teeth than does H. quelpaertensis (18.3 ± 0.98 (13 – 20) in males and 19.9 ± 1.42 (18 – 23) in females of the new species versus 23.8 ± 3.06 (18 – 30) in males and 22.8 ± 1.25 (21 – 24) in females of H. quelpaertensis). The new species can be further diagnosed from H. leechii from mainland Korea by its relatively shorter tail length: TL / SVL ratio is 0.79 ± 0.01 (0.54 – 0.98) in males and 0.73 ± 0.02 (0.55 – 0.89) in females of the new species versus 0.89 ± 0.05 (0.76 – 0.99) in males and 0.86 ± 0.03 (0.74 – 1.06) in females of H. leechii (TL, one-way ANOVA, df = 3, F = 9.360, P <0.01). H. unisacculus sp. nov. has a significantly shorter head, too: HL / SVL ratio is 0.24 ± 0.00 (0.21 – 0.26) in males and 0.23 ± 0.00 (0.21 – 0.25) in females of the new species versus 0.27 ± 0.01 (0.26 – 0.29) in males and 0.25 ± 0.00 (0.22 – 0.28) in females of H. leechii (HL, one-way ANOVA, df = 3, F = 9.356, P <0.01). The new species has a significantly longer trunk than does H. leechii: TRL / SVL ratio is 0.77 ± 0.00 (0.74 – 0.82) in males and 0.78 ± 0.00 (0.75 – 0.81) in females of H. unisacculus sp. nov. versus 0.73 ± 0.01 (0.71 – 0.74) in males and 0.75 ± 0.00 (0.72 – 0.78) in females of H. leechii (TRL, one-way ANOVA, df = 3, F = 7.514, P <0.01); in the new species when limbs are adpressed to the body, digit tips are always separated by a large gap comprising - 2.4 ± 0.42 (- 3 – - 1.5) of costal folds in males of H. unisacculus sp. nov. whereas there is only a small gap — less than one costal fold — in H. leechii (- 0.4 ± 0.35 (- 1 – 0 )). The new species is not significantly different in size and most body ratios from H. yangi, but H. unisacculus sp. nov. has slightly deeper vomerine tooth series than H. yangi (VTW / VTL in males of the new species 1.53 (1.00 – 1.78) versus 1.62 (1.24 – 1.89) in males of H. yangi); in the latter species vomerine series are slightly less curved than in the new species. Finally, H. unisacculus sp. nov. has relatively shorter limbs and a comparatively longer body than H. yangi: between adpressed limbs in the new species there is always a wide gap of - 2.4 costal folds in males and - 2.5 costal folds in females (ranging from - 3.5 – - 1.5), whereas in H. yangi males the adpressed limbs often in touch or overlap and the CGBL value is - 0.9 ± 0.32 (ranging from - 1.5 – - 0.5). The new species is also morphologically distinct from the undescribed sister lineage Hynobius clade 1 (HC 1) from coastal and lowland areas in the southern part of Gyeongsangnam-do Province in number of morphological traits: in having 11 – 12 costal folds (11.1 ± 0.18) in males (versus 10 costal folds in males of Hynobius clade 1); in having much shorter tail, TL / SVL ratio is 0.74 ± 0.18 in males (versus longer tail in males of Hynobius clade 1, TL / SVL ratio is 0.92 ± 0.10); in having relatively shorter hindlimbs, HLL / SVL ratio in males of the new species is 0.26 ± 0.01 (versus comparatively longer hindlimbs, HLL / SVL ratio is 0.26 ± 0.01 in males of Hynobius clade 1); by comparatively smaller head, in the males of the new species HL / SVL ratio is 0.24 ± 0.01 and HW / SVL ratio is 0.16 ± 0.00 (vs. comparatively larger head in males of Hynobius clade 1: HL / SVL ratio is 0.27 ± 0.01 and HW / SVL ratio is 0.18 ± 0.00); by comparatively shorter eye to snout distance, OR / SVL ratio is 0.07 ± 0.00 in males of the new species (versus OR / SVL ratio is 0.09 ± 0.00 in males of Hynobius clade 1); finally, between adpressed limbs in the new species there is always a wide gap of - 2.4 costal folds in males (ranging from - 3.0 – - 1.5), whereas in Hynobius clade 1 males the adpressed limbs usually in touch or overlap and the CGBL value is 0.0 ± 0.2 (ranging from - 1.0 – 1.0). Other lentic-breeding Hynobius species found in East and Northeast Asia can be distinguished from H. unisacculus sp. nov. by a combination of several morphological characters. H. tsuensis Abé, inhabiting the Tsushima Islands in the Korea Strait, can be distinguished from the new species by much longer vomerine tooth series (VTW / VTL ratio 1.13 (0.98 – 1.27) versus 1.53 (1.00 – 1.82) in the new species; data for males), by a longer tail (TL / SVL ratio 0.82 (0.71 – 0.86) versus 0.79 ± 0.01 (0.54 – 0.98) in the new species; data for males) and by a bright yellow stripe on the dorsal surface of the tail (versus absent in the new species). H. nebulosus (Temminck et Schlegel), inhabiting Kyushu and the western part of Honshu Island in Japan, also has bright-yellow stripe on the dorsal edge of the tail (versus absent in H. unisacculus sp. nov.), deeper vomerine tooth series (VTW / VTL ratio 1.04 (0.85 – 1.32) versus 1.53 (1.00 – 1.82) in the new species; data for males), and a longer tail (TL / SVL ratio 70.5 (62.2 – 80.4) versus 0.79 ± 0.01 (0.54 – 0.98) in the new species; data for males). H. dunni Tago, from the western part of Kyushu and the easternmost Shikoku, can be distinguished from H. unisacculus sp. nov. by deeper vomerine series; longer limbs, overlapping or in touch when adpressed to body; uniform brown-olive coloration (versus shallower vomerine series; smaller limbs, separated by a wide gap of 3 costal folds; bronze-spotted darkbrown coloration in the new species). Several Japanese lentic-breeding species can be distinguished from the new species by coloration; they all have numerous silvery, whitish or bluish spots and speckles on the dorsum and body flanks: H. lichenatus Boulenger from northeastern Japan (Tohoku, Honshu); H. takedai Matsui et Miyazaki from the Hokuriku region of central Honshu, along the coast of the Sea of Japan (Eastern Sea); and H. hidamontanus Matsui from mountain areas in central Honshu. H. tokyoensis Tago from the Kanto area of eastern Honshu usually lacks bluish spots, but has more costal grooves than does the new species: from 12 to 14 costal grooves (median — 13 costal grooves) in males. H. abei Sato from the Sea of Japan (Eastern Sea) coast of western Honshu differs from H. unisacculus sp. nov. in its longer and broader head, shorter trunk, and much higher tail. The above-mentioned species can be further diagnosed from the new species by its comparatively longer limbs with digits overlapping or separated by a small gap when limbs are adpressed to the body; in males, the CGBL value varies from 0 to 2.5 in H. lichenatus, from - 1 to 1 in H. takedai, from - 2 to 1 in H. tokyoensis, from - 2 to - 0.5 (mean - 2) in H. abei and from - 2 to - 0.5 (mean - 1) in H. hidamontanus. H. nigrescens Stejneger inhabiting northeastern Japan (Tohoku, Honshu), can be easily diagnosed from H. unisacculus sp. nov. by its larger body size (total length up to 190 mm; SVL = 66.4 ± 0.94 (60.5 – 73.4 mm) in males), longer tail and longer limbs, which overlap on 1 - 3 costal segments when adpressed to the body (versus smaller size and shorter limbs, separated when adpressed to body by never less than 3 costal segments in the new species). H. unisacculus sp. nov. can be diagnosed from the group of Chinese lentic-breeding Hynobius, which inhabit eastern and central parts of China, by its smaller body size (SVL up to 61 mm; mean SVL 49.0 mm in males and 51.3 mm in females): versus H. amjiensis Gu (SVL in males 83.6, from 79.5 to 86.5), H. guabangshanensis Shen (SVL in males 81.4, from 72.8 to 87.5), H. maoershanensis Zhou, Jiang et Jiang (SVL in males 86.1, from 83.1 to 91.1), and H. yiwuensis Cai (SVL in males 63.8, from 47.0 to 74.0). H. chinensis Günther, H. amjiensis and H. maoershanensis also have tails longer than the new species (mean TL / SVL values 0.85, 0.92 and 0.84, respectively). The new species markedly differs from all other Hynobius in its breeding biology; to date in all types of waterbodies used for breeding that were examined by us we observed only single egg sacs, which lacked the gelatinous stalk-like structure typical for other lentic-breeding Hynobius (Song & Koo 2010). Finally, the new species is markedly distinct from all other congeners for which comparable sequences are available, including its closest relatives from the Korean Peninsula in its large genetic distances in the 12 S rRNA mtDNA (genetic distance from p = 9.40 %) and COI gene (genetic distance from p = 9.10 %) fragments (see Table 4). This degree of pairwise divergence in the 12 S rRNA and COI gene is greater than that usually representing differentiation at the species level in amphibians (Smith et al. 2008; Xia et al. 2012; Murphy et al. 2013). To date, despite intensive fieldwork in the southern part of the Korean Peninsula we have not found any signs of sympatry of the new species with other Korean Hynobius (see Baek et al. 2011 a, b). Phylogenetic position. Baek et al. (2011 a, b) compared mitochondrial DNA sequences between the new species and the Korean hynobiids Hynobius leechii, H. quelpaertensis, and H. yangi. The analysis of the partial cytochrome b gene (907 bp), COI (1,097 bp), and the 12 S rRNA (788 bp) gene showed profound differences between the new species and other Korean Hynobius (see Table 5 and Results for details). The observed level of genetic divergence between the new species and other Korean Hynobius clearly corresponds to a species level of differentiation (Lee et al. 1998; Lai & Lue 2008). Based on the data of Baek et al. (2011 a, b), H. unisacculus sp. nov. is recovered as a sister species of lineage HC 1, both lineages forming a clade with H. quelpaertensis.
Min, Mi-Sook, Baek, Hae-Jun, Song, Jae-Young, Chang, Min Ho, Poyarkov Jr, Nikolay A. (2016): A new species of salamander of the genus Hynobius (Amphibia, Caudata, Hynobiidae) from South Korea. Zootaxa 4169 (3): 475-503, DOI: 10.11646/zootaxa.4169.3.4