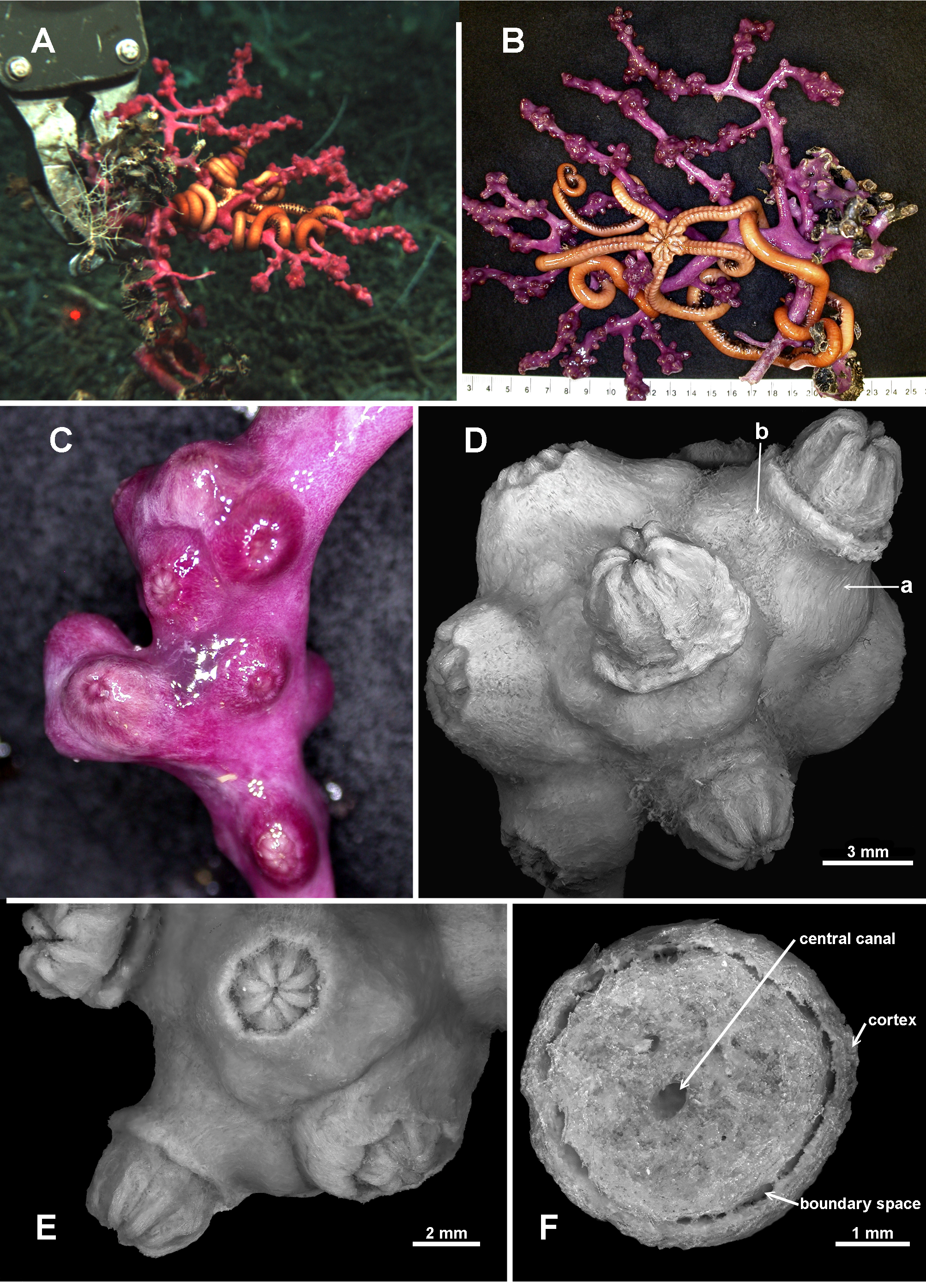
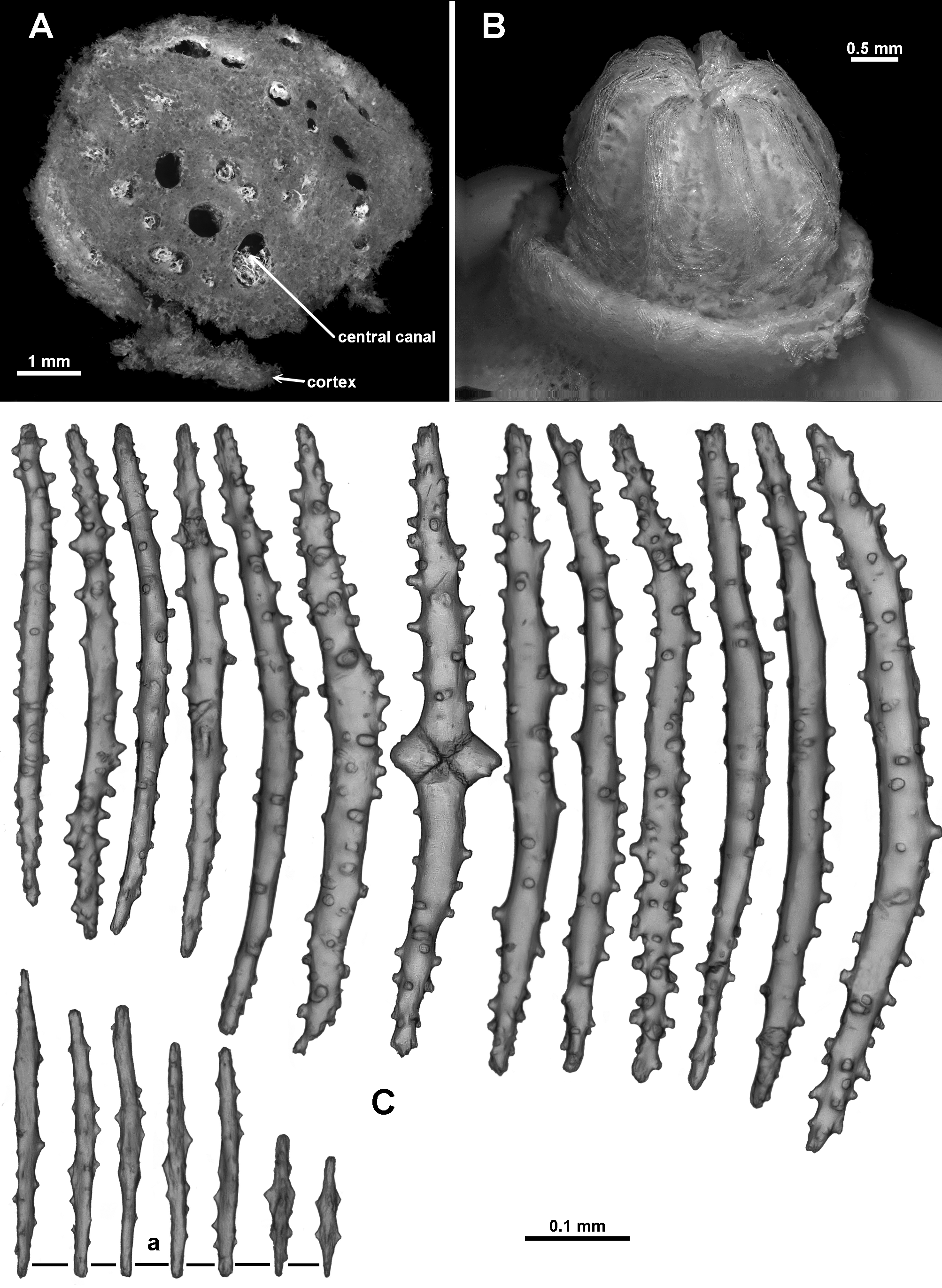
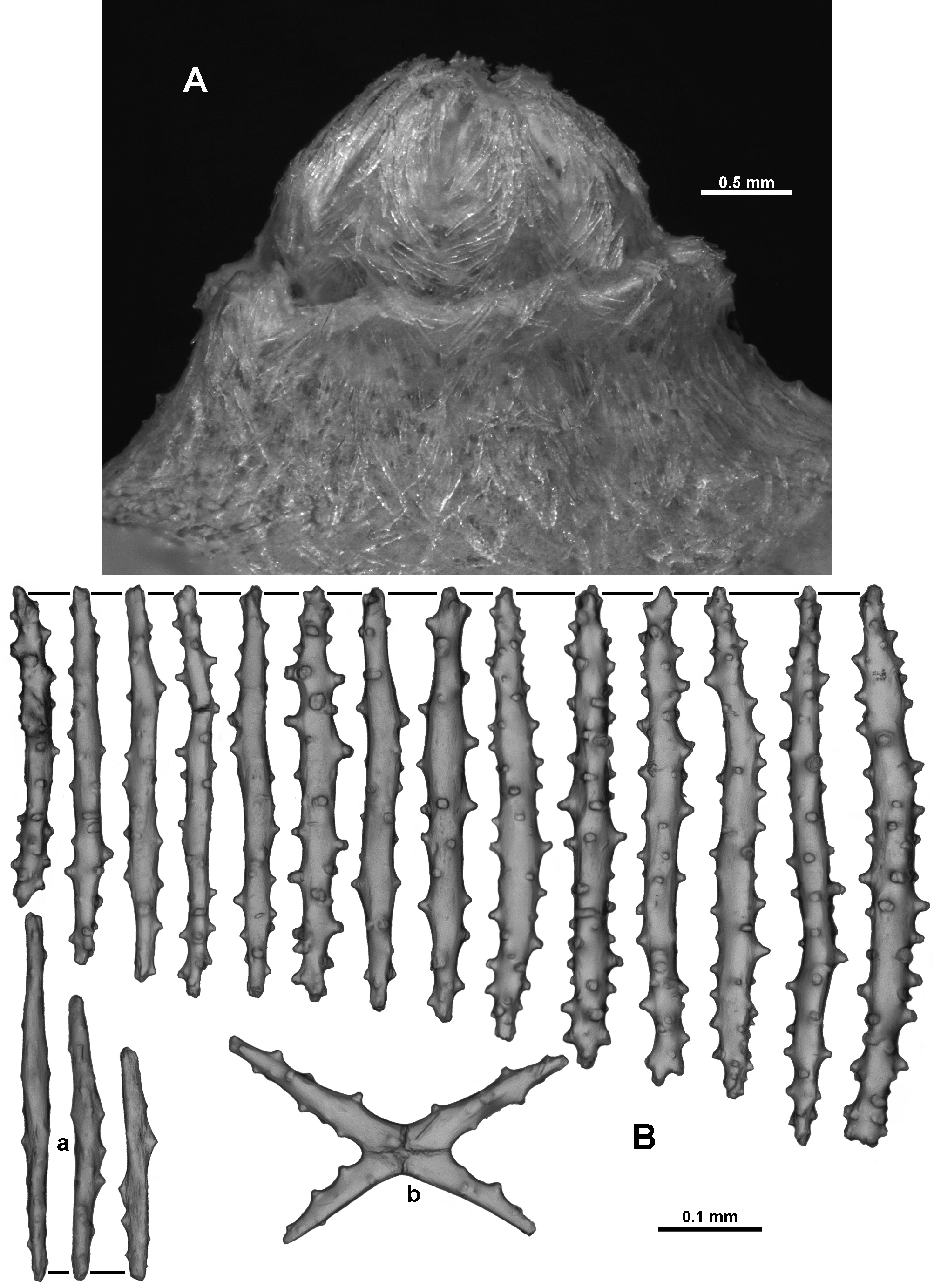
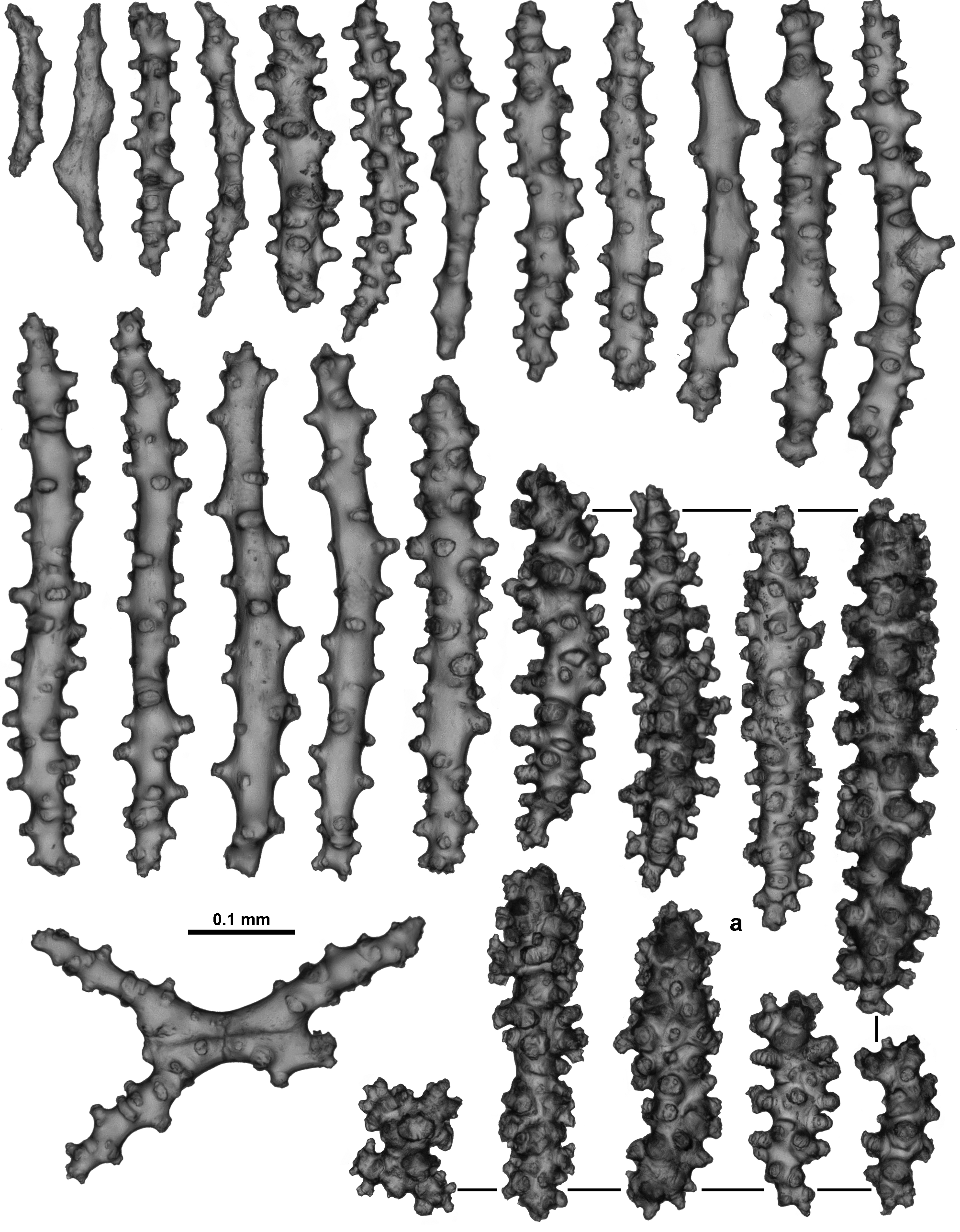
Material examined. Holotype: TMAG K 4266, Z 27 Seamount, Huon Commonwealth Marine Reserve (CMR), SW Tasman Sea, Australia, stn. J 2 - 385 - 005, sample 010 b, 44.245 ° S, 147.121 ° E, depth 1060 m, ROV Jason deployed from the U. S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 21 st December 2008. Paratypes: TMAG K 4267, Z 27 Seamount, Huon CMR, SW Tasman Sea, Australia, stn. J 2 - 385 - 005, sample 010 a, 44.245 ° S, 147.121 ° E, depth 1060 m, ROV Jason deployed from the U. S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 21 st December 2008; TMAG K 4268, Mongrel Seamount, Huon CMR, SW Tasman Sea, Australia, stn. J 2 - 386 - 006, sample 0 0 1, 44.254 ° S, 147.115 ° E, depth 982 m, ROV Jason deployed from the U. S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23 rd December 2008; TMAG K 4269, Mongrel Seamount, Huon CMR, SW Tasman Sea, Australia, stn. J 2 - 386 - 011, sample 0 21, 44.255 ° S, 147.114 ° E, depth 899 m, ROV Jason deployed from the U. S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23 rd December 2008; TMAG K 4270, Mongrel Seamount, Huon CMR, SW Tasman Sea, Australia, stn. J 2 - 386 - 007, sample 0 0 3, 44.254 ° S, 147.114 ° E, depth 958 m, ROV Jason deployed from the U. S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23 rd December 2008; TMAG K 4115, Hill V Seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 69, (SS 199701 / 69), 44.397 – 44.398 ° S, 147.147 – 147.178 ° E, depth 1262 – 1854 m, 31 st January 1997; NTM CO 13052 (ex TMAG K 1360), Hill J 1 Seamount, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 36, (SS 199701 / 36), 44.267 – 44.242 ° S, 147.332 – 147.363 ° E, depth 1518.4 m, 27 th January 1997; TMAG K 4271, E. N. E. of St. Patricks Head, eastern Tasmania, Australia, CSIRO RV Soela, stn. 16, (SO 198705 / 16), 41.573 S, 148.743 E, depth 1090 – 1150 m, 9 th May 1987. Other material: NTM CO 13050 (ex TMAG K 1362), Dory Hill Seamount, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 47, (SS 199701 / 47), 44.322 – 44.34 ° S, 147.115 – 147.072 ° E, depth 1280 – 1400 m, 29 th January 1997.
Moore, Kirrily M., Alderslade, Philip, Miller, Karen J. (2017): A taxonomic revision of Anthothela (Octocorallia: Scleraxonia: Anthothelidae) and related genera, with the addition of new taxa, using morphological and molecular data. Zootaxa 4304 (1): 1-212, DOI: 10.11646/zootaxa.4304.1.1