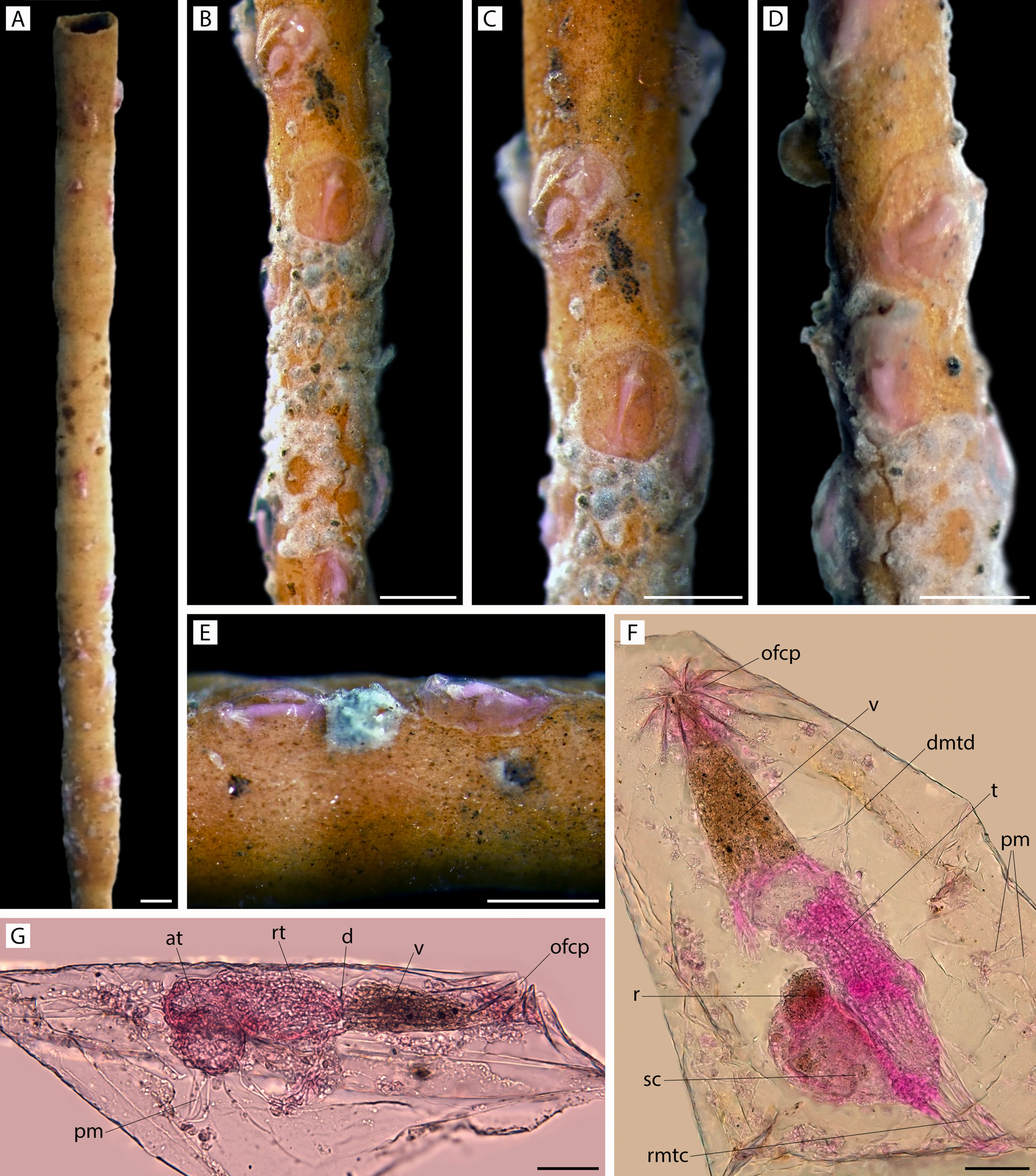
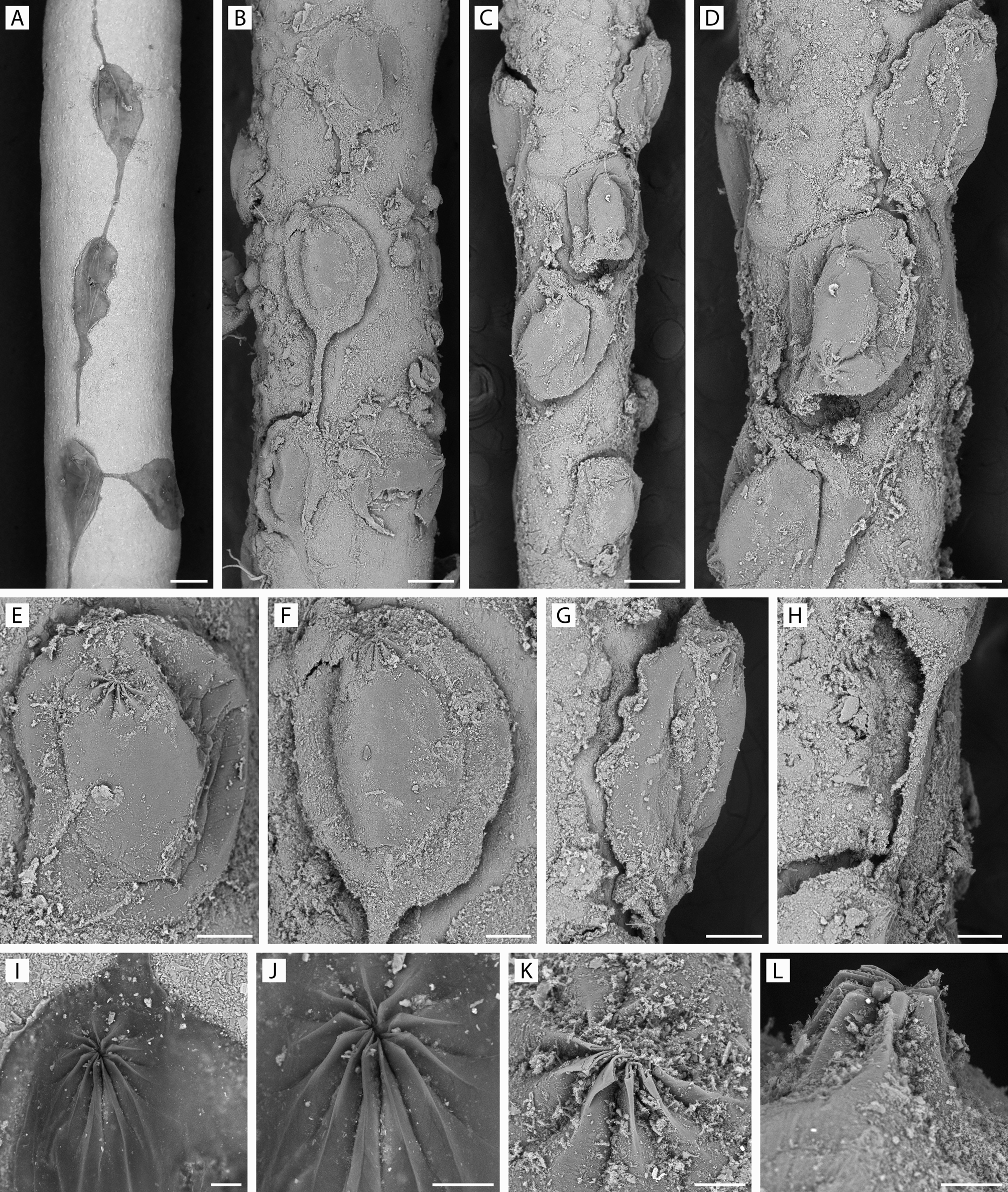
Remarks. We include this new species in the otherwise monotypic genus Pierrella, previously known only as a fossil taxon from the Cretaceous (lower Campanian to lower Maastrichtian) of Wyoming and South Dakota, USA. The type species, P. larsoni, was preserved on inner shell surfaces of empty body chambers of ammonites by a process interpreted as lithoimmuration. The overall characters of P. larsoni accord with living members of Arachnidiidae in terms of colony form (adnate, uniserial, branching) and zooid shape (clavate / caudate), but the genus was able to be distinguished by its distinctive pleats (folds), eight in all, forming the orifice in the retracted state. This arrangement is unknown in any other arachnidioid, in which the orifice, when closed, has been described as a mamilla or papilla, or is somewhat elevated and squared (d'Hondt 1983). Wilson & Taylor (2013) referred to the pleats in P. larsoni as " setigerous, or pleated, collars ", but it remains to be seen if the stiff orificial folds in Pierrella are homologous with the setigerous apparatus of the introvert in vesicularioid ctenostomes. For example, in the ctenostomes studied by McKinney & Dewel (2002), all collars were " composed of membranes that appeared to have their basal attachment at the atrial sphincter (diaphragm) "; these authors distinguished four forms of collar. Notwithstanding this uncertainty, which has no bearing on the relatedness of Pierrella plicata n. sp. to P. larsoni, the similarity of these two species is compelling. They accord in every important respect — colony form, zooid shape, zooid size, and the overlapping number of stiff orificial pleats (eight in P. larsoni, 8 – 11 in P. plicata). The only differences concern budding locus (somewhat more distolateral in P. larsoni) and budding pattern (morevariable branch angles in P. larsoni), but these differences can be intrageneric. In both species, zooid length and shape vary, becoming longer and more clavate in distal colony parts. In P. plicata, a squatter zooid shape occurs where zooids are more crowded. An apparent colony origin was found in P. larsoni, giving evidence of a nearcircular ancestrula with two distolateral buds and a proximal bud. An ancestrula was not seen in P. plicata; its discovery would give more information concerning relatedness. The long temporal gap of c. 74 million years need not preclude these two species being congeneric. A perusal of the Bryozoa Home Page shows that there are quite a number of Mesozoic genera, from three orders, still present in modern seas, e. g. Arachnidium, Arachnoidella, Buskia (Ctenostomata), Nellia, Onychocella, Poricellaria, Celleporella (Cheilostomata), Microeciella, Plagioecia, Mesenteripora, Crisia (Cyclostomata) (see also Voigt 1985). Pierrella plicata n. sp. encrusts tubular arenaceous foraminiferans resembling Rhabdammina, which live at the sediment surface. The largest colonies comprised fewer than ten zooids. The proportionately very long vestibule in zooids examined in transparency and stained with Rose Bengal give evidence that, when the polypide retracts, the vestibule fills with fine sediment particles. These would be emptied from the vestibule upon the next polypide eversion.
Grischenko, Andrei V., Gordon, Dennis P., Melnik, Viacheslav P. (2018): Bryozoa (Cyclostomata and Ctenostomata) from polymetallic nodules in the Russian exploration area, Clarion - Clipperton Fracture Zone, eastern Pacific Ocean-taxon novelty and implications of mining. Zootaxa 4484 (1): 1-91, DOI: 10.11646/zootaxa.4484.1.1