Rallocytus ridiculus
Grischenko, Gordon & Melnik, 2018
GBIF:148403734
ABOUT
Descriptions(5)
Export occurrence data
Darwin Core Archive (ZIP)
CLASSIFICATION
Taxonomic Classification Tree

MULTIMEDIA
Media Files(5)
FIGURE 5. Pandanipora helix n. gen., n. sp. A, K, specimen YMG4–13, Stn 289; B, C, specimen YMG4–13, Stn 308; D, specimen YMG4–07, Stn 143; E, specimen GLD4–11, Stn 219; F, specimen YMG4–14, Stn 330; G–I, specimen YMG4–14, Stn 326; J, specimen GLD4–12, Stn 258; L, specimen YMG4–13, Stn 321. A–D, F, prop morphology: A, vertical and oblique columnar props attached to nodule by expanded tips; B, curved props attached to nodule irregularities by expanded tips with complex outlines; C, same, close-up; D, single elongated prop from its inception in the zooid basal wall to its distal expansion; E, light micrograph of colony showing successively overlapping zooids with their coelomic cavities continuing into the props with no intervening septum or pore, as well as 2–5 brown bodies located centrally to proximally in several zooids; F, prop tip showing opening towards substratum and internal surface with wedge-shaped crystallites, structurally analogous to peristome; G–I, ancestrula and daughter zooid budded from peristome; J–L, ancestrula and postancestrular zooids. Scale bars: A, B, E, 500 µm; C, D, J–L, 250 µm; F, 50 µm; G–I, 100 µm.
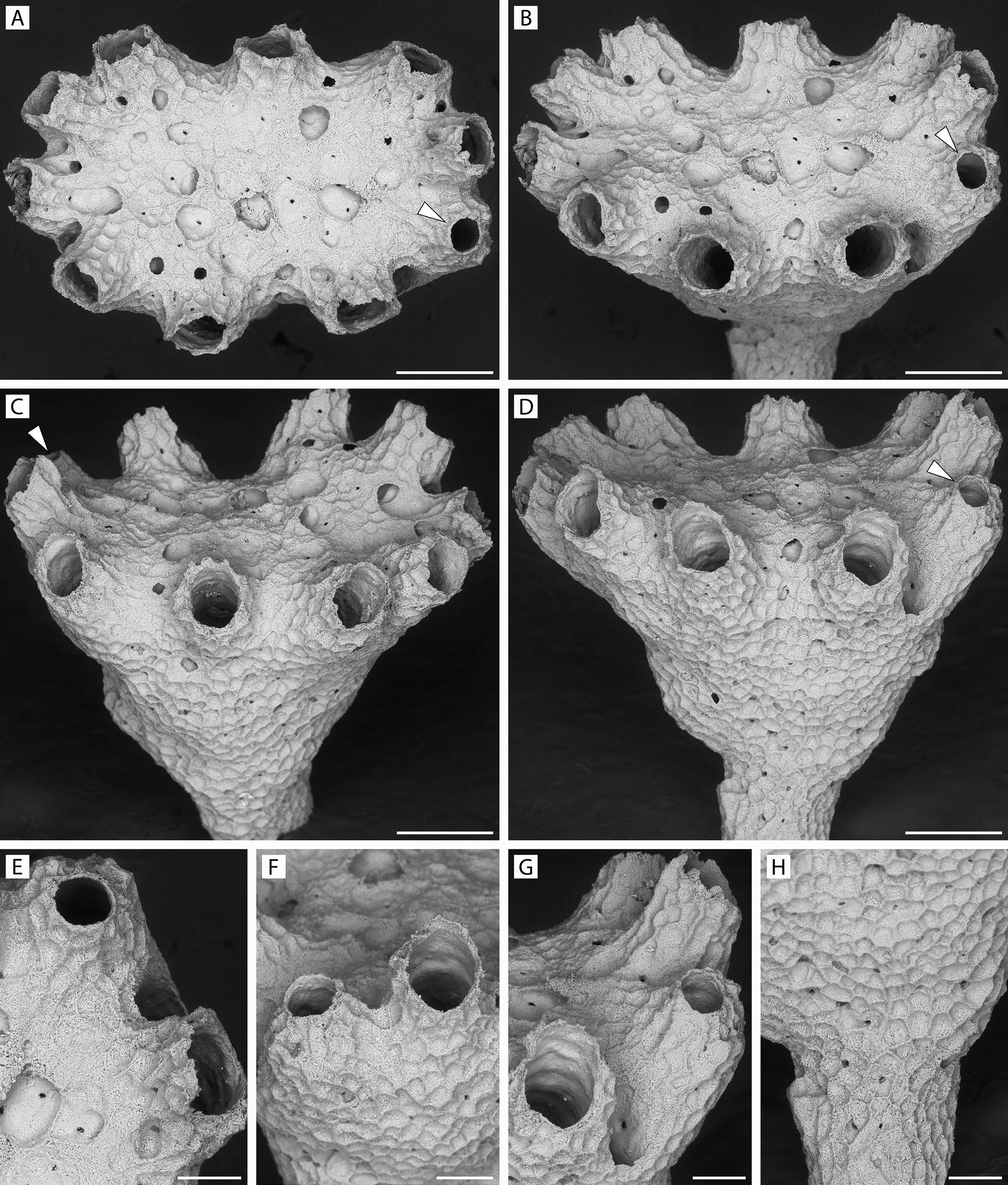
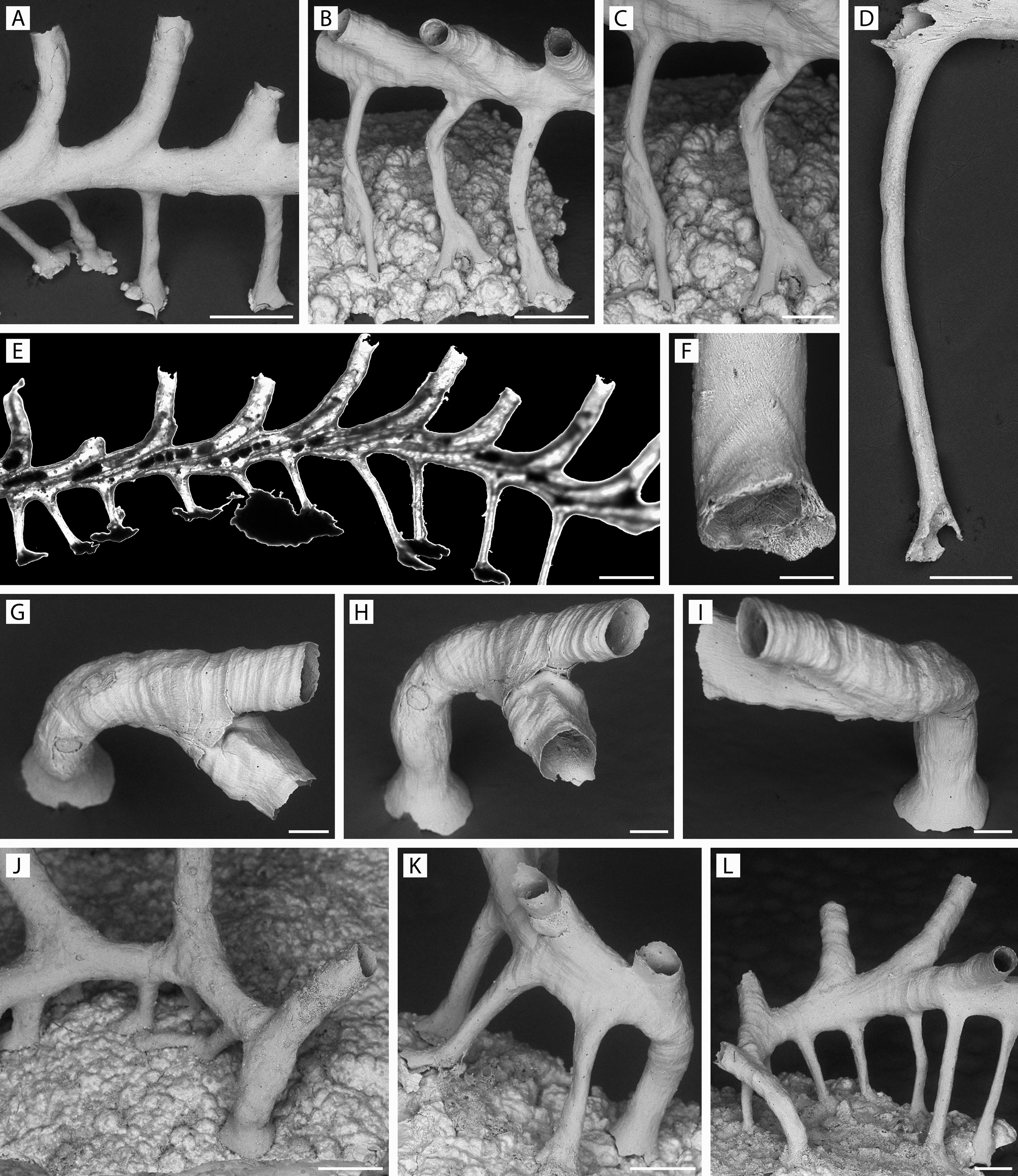
FIGURE 42. Rallocytus ridiculus n. gen., n. sp. Holotype, ZIRAS 1/50723. A–D, apical and oblique-lateral rotational views of colony with single dimorphic zooid, its smaller aperture (arrowed) facing frontalwards; E–G, peristomes and apertures of dimorphic zooid and adjacent autozooids; H, malleated colony surface at junction of capitulum and column. Scale bars: A–D, 250 µm; E–H, 100 µm.

FIGURE 43. Rallocytus ridiculus n. gen., n. sp. Paratype 1, ZIRAS 2/50724. A–D, apical and oblique views of fertile colony with two dimorphic zooids (arrowed), one with its aperture facing frontalwards, the other peristome turned slightly toward calyx center; E, F, peristomes, including a smaller frontally facing dimorphic one; G, two autozooidal peristomes with incurved dimorphic peristome between; H, base of column; I, skeletal microstructure; J, K, showing dimorphic and autozooidal apertures, respectively. Scale bars: A–C, 250 µm; D–H, 100 µm; J, K, 50 µm; I, 25 µm.
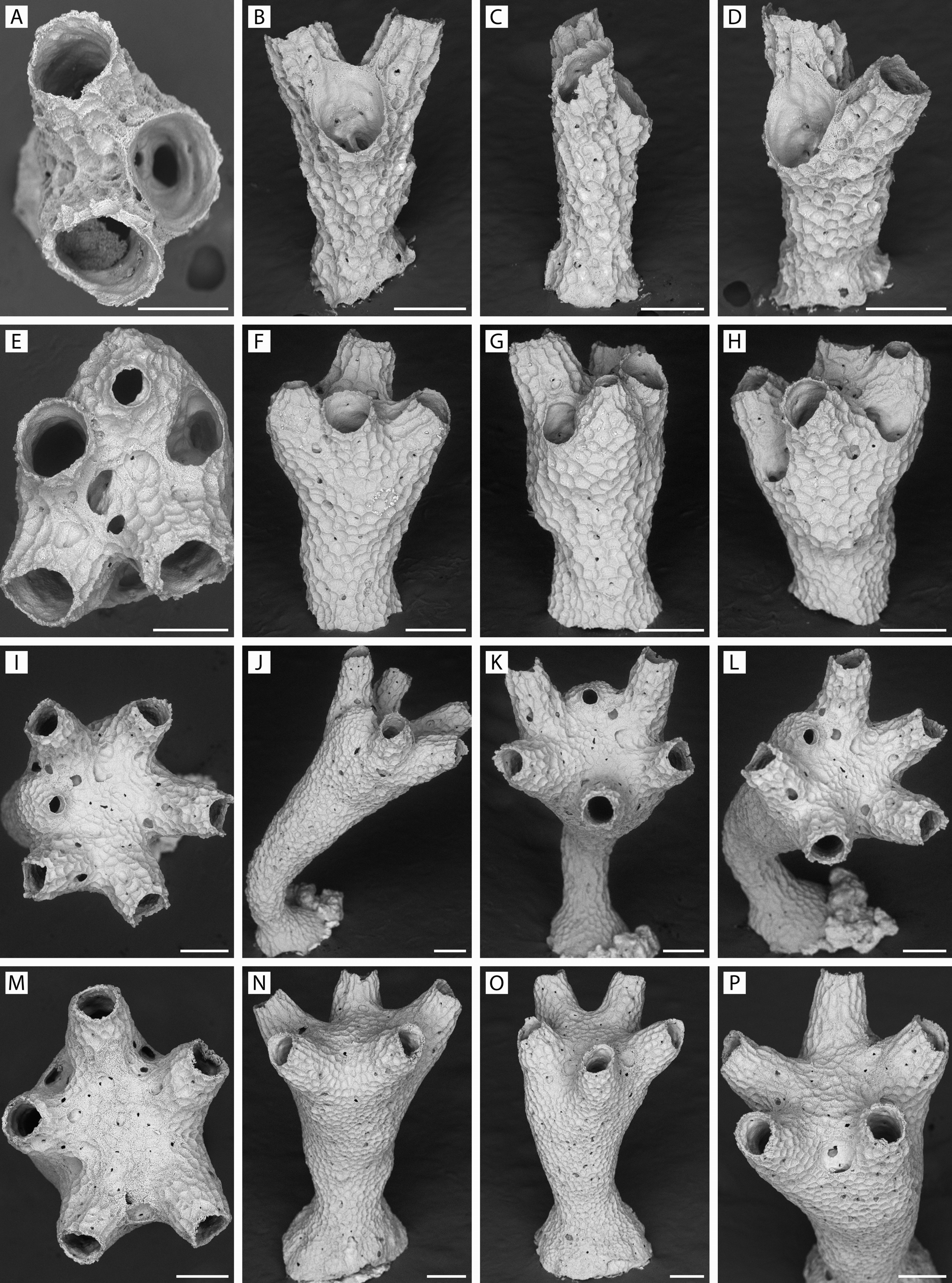
FIGURE 44. Rallocytus ridiculus n. gen., n. sp. Progressive stages of development of ancestrulate colonies. A–D, specimen YMG4–14, Stn 340, at three-zooid stage, with no distinction among peristomes; E–H, specimen GLD4–09, Stn 183, with four functional zooids, one dimorphic; I–L, specimen GLD4–08, Stn 159, with six functional zooids, one dimorphic; M–P, specimen GLD4–11, Stn 206, five functional zooids, none dimorphic. Scale bars: A, E, 150 µm; B–D, F–P, 200 µm.
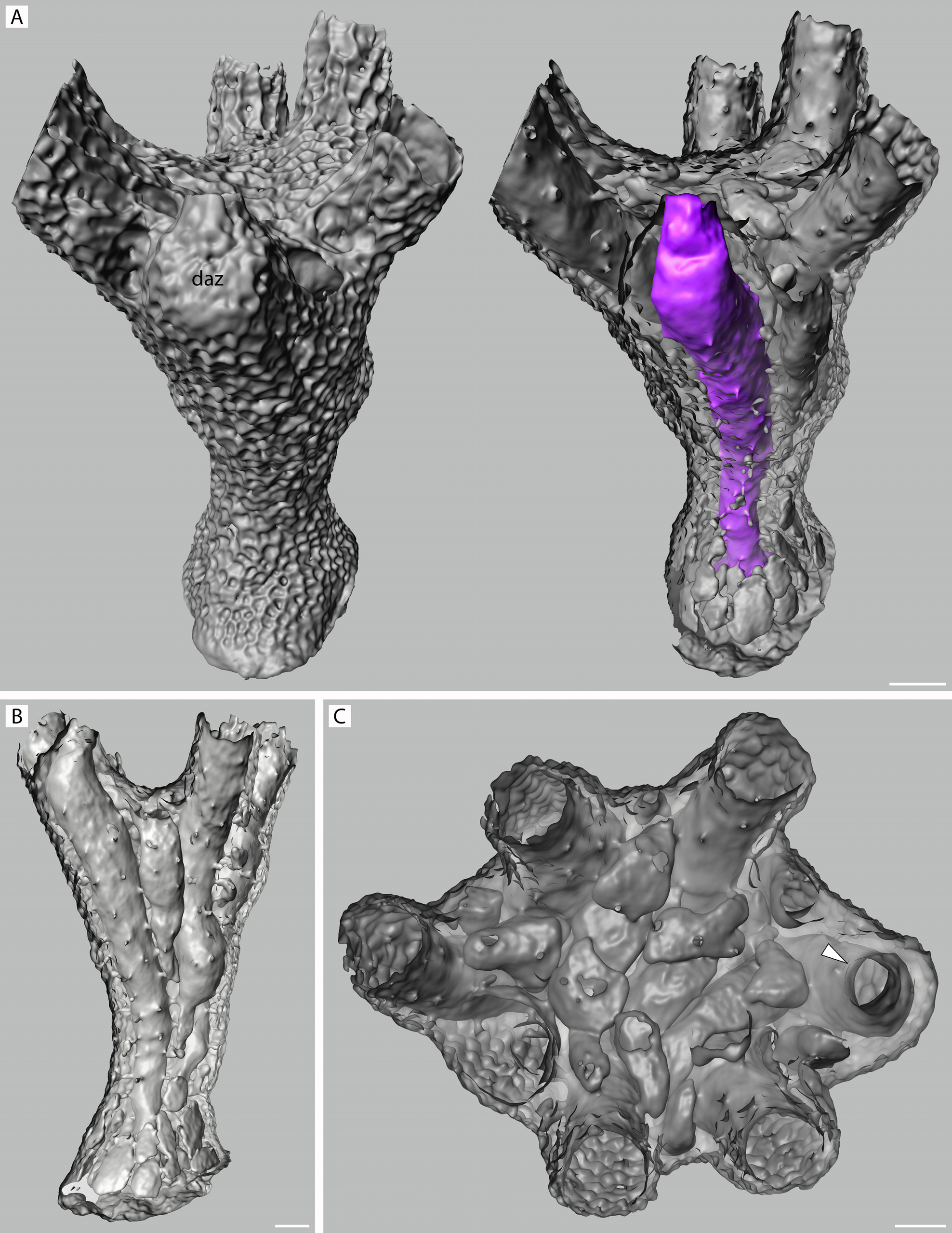
FIGURE 54. Rallocytus ridiculus n. gen., n. sp. Micro-CT scans of paratype 2, NIWA 127726, as surface (A) and back-face isosurface renders showing colony interiors. A, surface and interior views of seven-zooid colony with dimorphic ancestrular zooid (daz) colored purple in the right-hand image; note small cluster of flattened kenozooidal chambers surrounding protoecium but absent from sides of column; B, lateral view, with entire dimorphic ancestrular zooid seen in left half; C, apical view of calyx, showing flattened kenozooidal/alveolar chambers filling center, dimorphic peristome at right indicated by arrowhead. Scale bars: 100 µm.
IMAGES