
Pandanipora helix
Grischenko, Gordon & Melnik, 2018
GBIF:148403742
ABOUT
Descriptions(6)
Export occurrence data
Darwin Core Archive (ZIP)
CLASSIFICATION
Taxonomic Classification Tree

MULTIMEDIA
Media Files(4)
FIGURE 2. Colonies of some cyclostome bryozoans, in vivo, attached to polymetallic nodules. A–E, Pandanipora helix n. gen., n. sp.: A, specimen GLD4–09, Stn 190; B, specimen GLD4–12, Stn 262; C, specimen YMG4–07, Stn 143; D, specimen YMG4–13, Stn 295; E, specimen GLD 4–11, Stn 212. F, Tubuliporina sp. indet., specimen YMG18–01, Stn 7. G, H, Abyssoecia elevata n. gen., n. sp.: G, specimen GLD4–09, Stn 196; H, specimen GLD4–09, Stn 191. I, Discantenna metallica n. sp.: specimen GLD4–11, Stn 224. J, K, Frontohornera frontalis n. gen., n. sp.: J, specimen YMG4–07, Stn 124; K, specimen GLD4–11, Stn 210. L, Alyonushka hystricosa n. gen., n. sp.: specimen GLD4–09, Stn 199. M, Calyssopora volcano n. gen., n. sp.: specimen YMG18–01, Stn 33. N, O, Anyuta anastema n. gen., n. sp.: N, specimen GLD4–09, Stn 180; O, specimen YMG4–06, Stn 71. Scale bars: 1 mm.
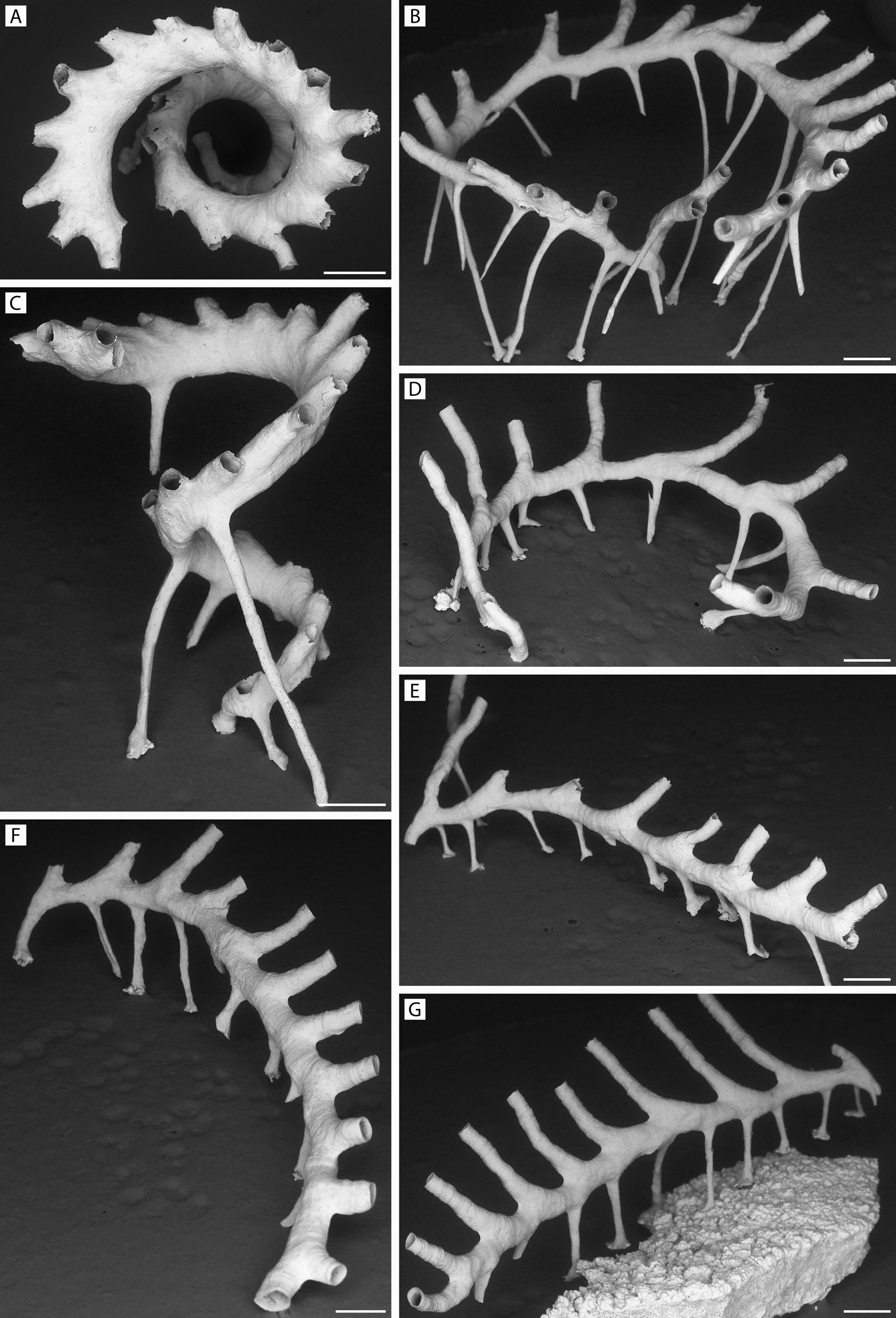
FIGURE 3. Pandanipora helix n. gen., n. sp. Holotype and paratype colonies. A, C, holotype, ZIRAS 1/50667; B, paratype 1, ZIRAS 2/50668; D, paratype 2, ZIRAS 3/50669; E, paratype 3, ZIRAS 4/50670; F, paratype 4, ZIRAS 5/50671; G, paratype 5, ZIRAS 6/50672. A, apical, and C, lateral views of the spiral holotype colony with widened apical part of axis of zooids with mostly broken, shortened peristomes and supported by rare, distanced, elongated tubular props; B, ring-like colony showing attenuated zooids with elongated, gently arcuate tubular peristomes tilted outwards, and supported by strongly elongated, slightly crooked filiform props; note one prop at lower left bifurcated near termination; D, crescentic colony showing zooids with elongated, arcuate tubular peristomes angled outwards, and supported by shortened straight props; E, colony bifurcated near point of origin showing linear chain of zooids with straight peristomes; supported by short props, some with widened terminations; F, arcuate colony with thicker zooids having widened tubular peristomes, supported by gradually shortening props; G, straight colony with greatly elongated peristomes, supported by regular series of props with expanded terminal bases, including four attached to piece of nodule. Scale bars: 500 µm.
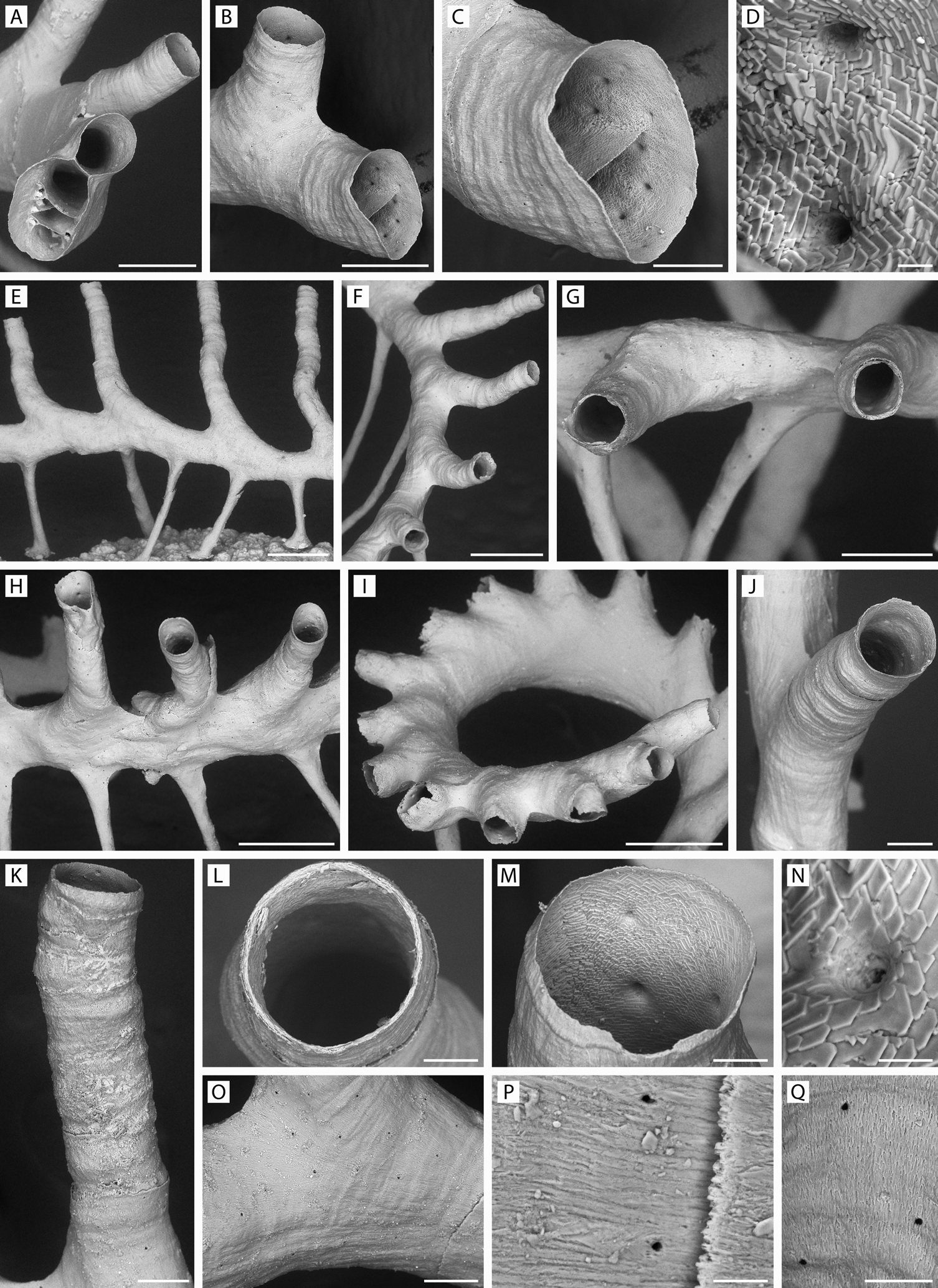
FIGURE 4. Pandanipora helix n. gen., n. sp. Details of morphology. A, specimen YMG4–13, Stn 289; B–D, paratype 4, ZIRAS 5/50671; E, paratype 5, ZIRAS 6/50672; F, G, Q, paratype 1, ZIRAS 2/50668; H, specimen GLD4–11, Stn 217; I, holotype, ZIRAS 1/50667; J, specimen YMG4–06, Stn 68; K, O, specimen YMG4–13, Stn 321; L, specimen YMG4–13, Stn 295; M, paratype 2, ZIRAS 3/50669; N, specimen GLD4–12, Stn 258; P, specimen GLD4–12, Stn 255. A, distal view of growing edge of colony showing early differentiation of daughter zooid and prop from the floor of the parent zooid, with chambers arranged in stacks; B, growing edge of colony showing differentiation of a daughter zooid (lower) from the floor of the parent zooid (upper); C, same, close-up; D, interior surface of developing zooid chamber showing imbricated foliated fabric of wedge-shaped crystallites and openings of pseudopores; E–I, variability of zooid size and geometry: E, elongated zooids with tall peristomes; F, part of ring-like colony; G, lateral view of same colony, two zooids near-perpendicular to plane of props; H, arcuate colony with overlapping peristome bases; I, lateral view of apical coil of spiral colony with subconnate peristome bases and shortened peristomes, most broken; J, frontolateral view of tubular peristome showing annular striae on the surface; K, lateral view of tubular peristome; L, peristomial aperture, frontal view; M, oblique view of peristomial aperture showing imbricated foliated fabric of wedge-shaped crystallites and interior openings of pseudopores; N, imbricated foliated fabric of wedge-shaped crystallites around internal opening of pseudopore; O, zooidal wall showing tiny sparse simple pseudopores; P, Q, close-up of zooidal wall showing crystallites. Scale bars: A, B, G, 250 µm; C, J, K, O, 100 µm; D, N, 10 µm; E, F, H, I, 500 µm; L, M, Q, 50 µm; P, 15 µm.
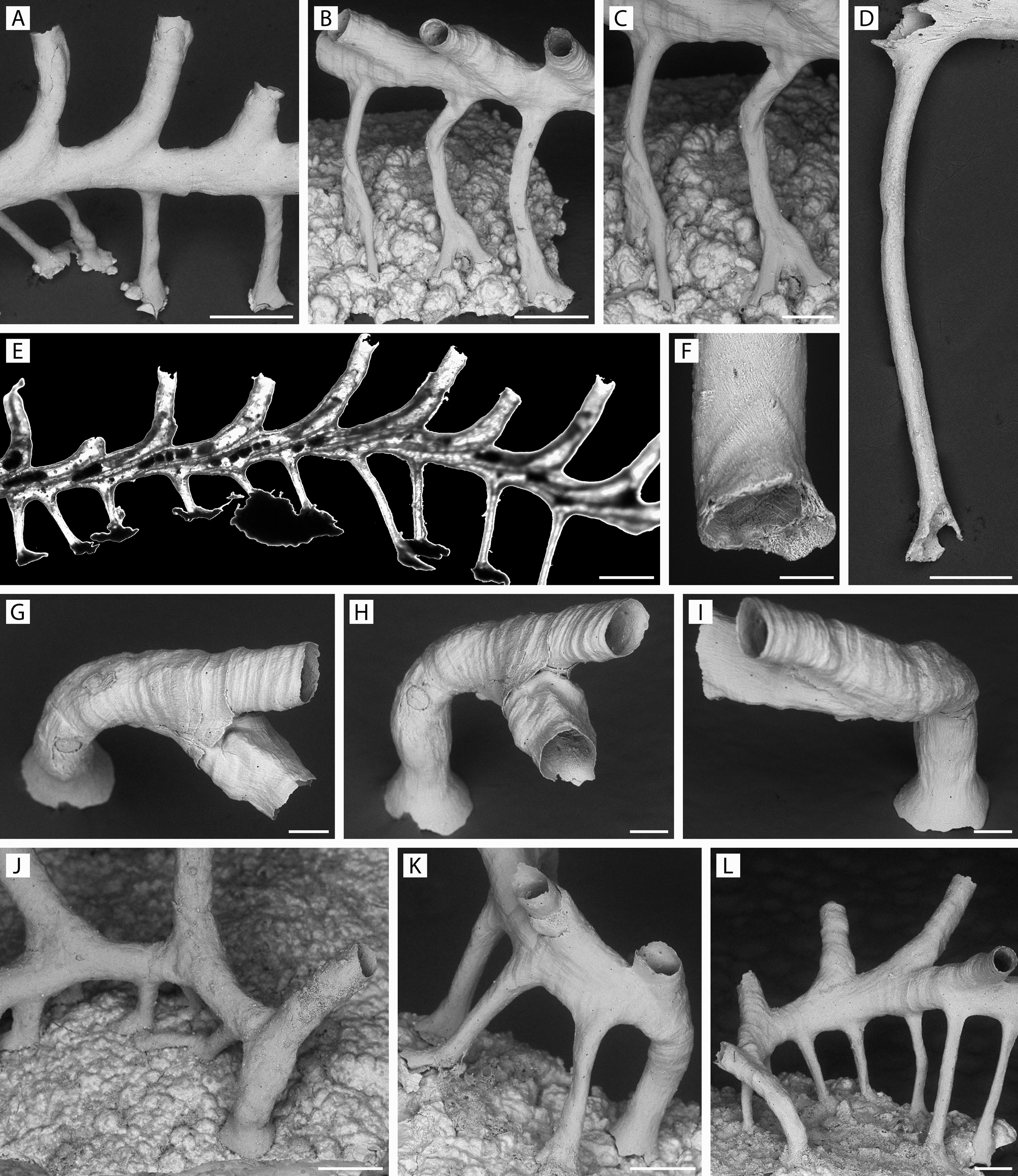
FIGURE 5. Pandanipora helix n. gen., n. sp. A, K, specimen YMG4–13, Stn 289; B, C, specimen YMG4–13, Stn 308; D, specimen YMG4–07, Stn 143; E, specimen GLD4–11, Stn 219; F, specimen YMG4–14, Stn 330; G–I, specimen YMG4–14, Stn 326; J, specimen GLD4–12, Stn 258; L, specimen YMG4–13, Stn 321. A–D, F, prop morphology: A, vertical and oblique columnar props attached to nodule by expanded tips; B, curved props attached to nodule irregularities by expanded tips with complex outlines; C, same, close-up; D, single elongated prop from its inception in the zooid basal wall to its distal expansion; E, light micrograph of colony showing successively overlapping zooids with their coelomic cavities continuing into the props with no intervening septum or pore, as well as 2–5 brown bodies located centrally to proximally in several zooids; F, prop tip showing opening towards substratum and internal surface with wedge-shaped crystallites, structurally analogous to peristome; G–I, ancestrula and daughter zooid budded from peristome; J–L, ancestrula and postancestrular zooids. Scale bars: A, B, E, 500 µm; C, D, J–L, 250 µm; F, 50 µm; G–I, 100 µm.
IMAGES