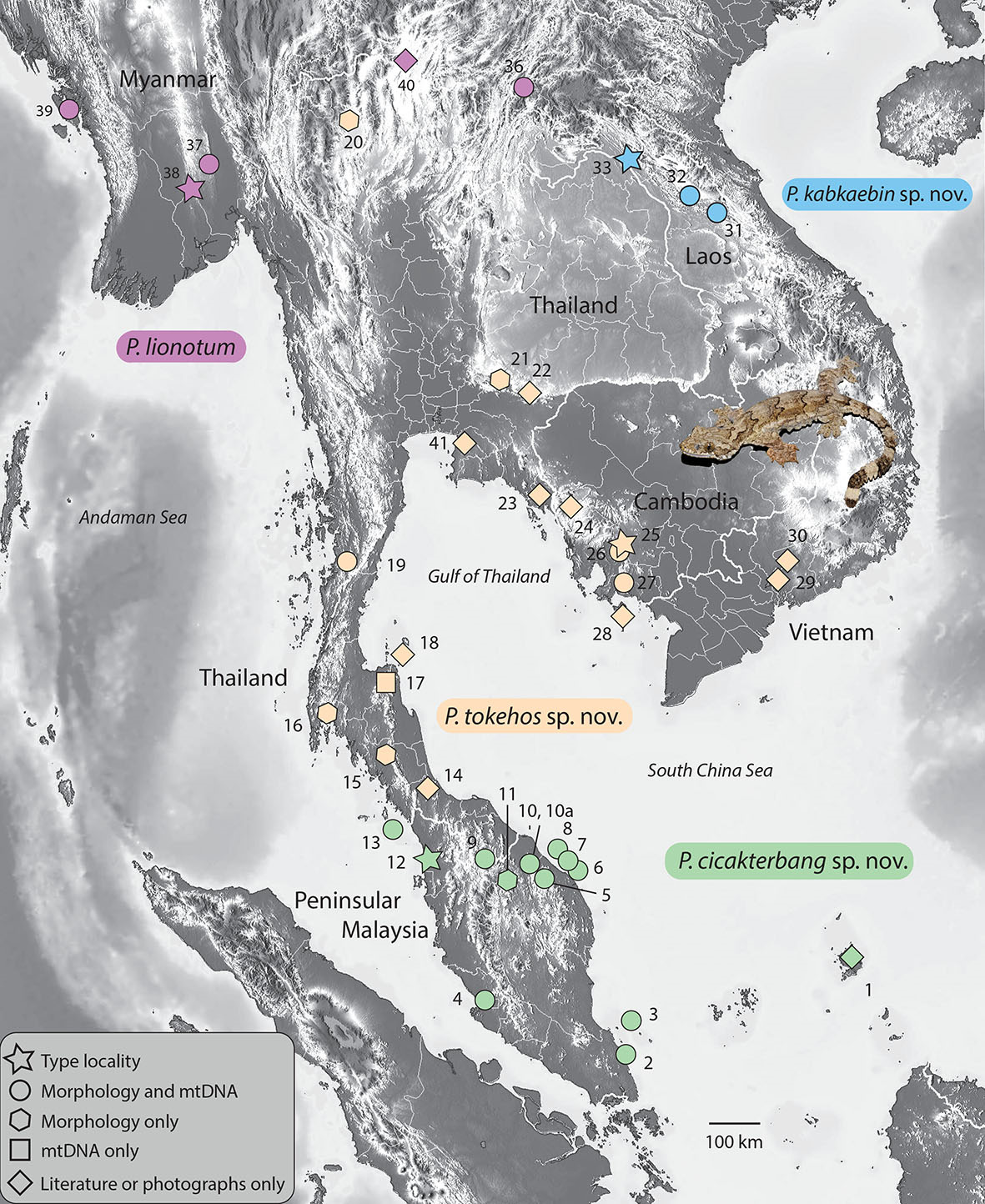
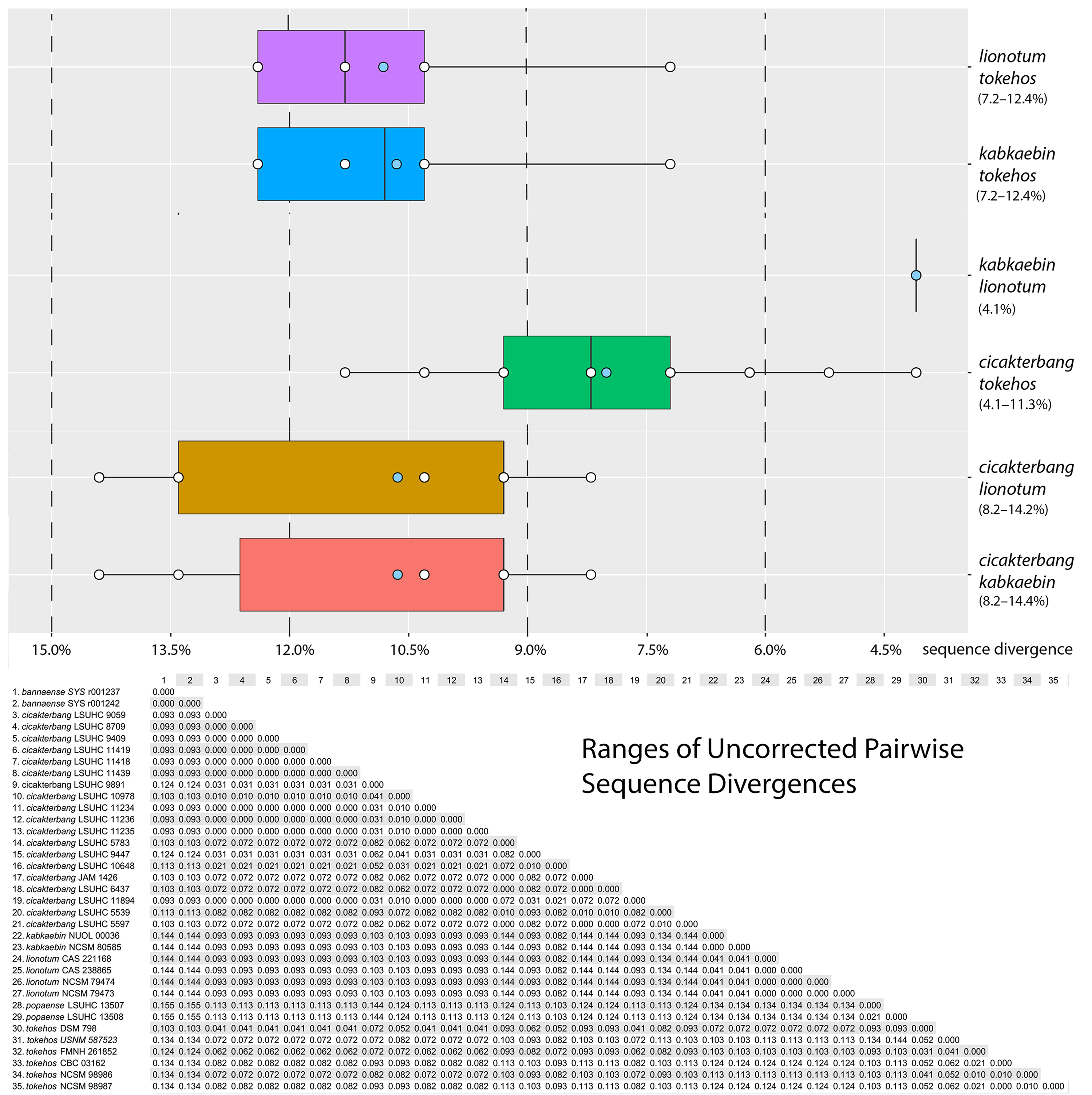
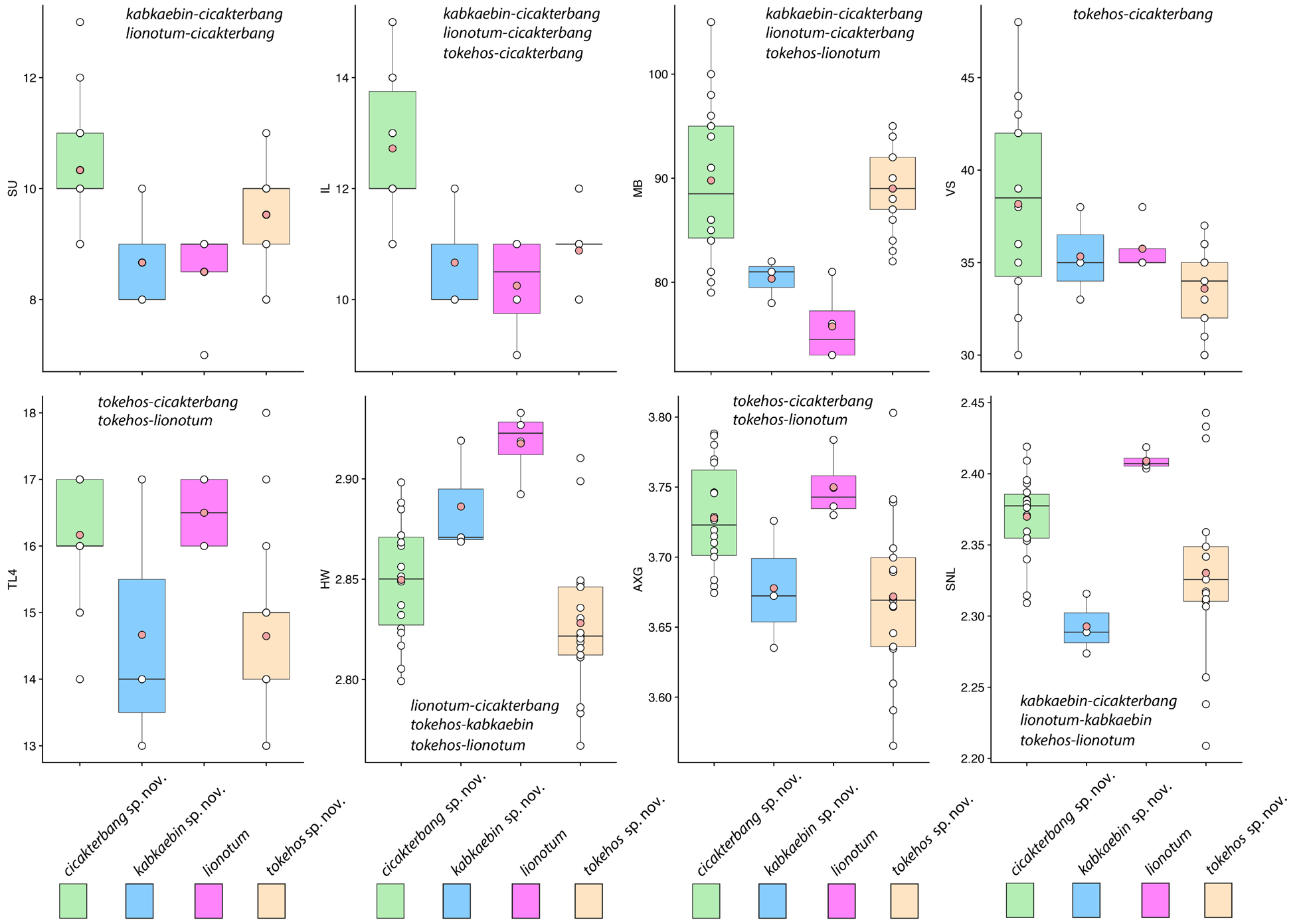
Description of holotype. Adult female SVL 86.0 mm; head moderate (HL / SVL 0.27), wide (HW / HL 0.76), depressed (HD / HL 0.40), distinct from neck; snout rounded at tip in dorsal profile; prefrontal region concave; lores rounded; rostral scale large, rectangular, bearing an inverted Y-shaped, dorsomedial groove, in contact posteriorly with two supranasals, one postnasal, dorsolaterally with nostrils, and laterally with first supralabials; supralabials (10 R, L) to mid-orbital position; infralabials (12 R, 14 L); nostrils elliptical with long axes oriented dorsoventrally, occupying anterior portion of nasal scale, bordered anteriorly by rostral, dorsally by supranasal, posteriorly by four postnasals of varying sizes (upper largest), and ventrally by first and second supralabials; scales on rostrum granular slightly larger than granular scales on top of head and occiput; eyes large (ED / HL 0.22), less than snout length; pupil vertically elliptical, crenelated; supraciliaries elongate, posteriormost weakly pointed; auricular opening rounded, bearing a prominent supra-auricular lobe; tympanum deeply sunk; infra-auricular flap broad, rounded, extending from below corner of mouth to lateral margin of neck midway between posterior margin of ear opening and forelimb insertion, measuring 3.9 mm at its widest point; dorsal scales of infra-auricular flap large, subimbricate proximally, small juxtaposed distally, ventral flap scales minute and granular; mental triangular, slightly wider than deep, bordered laterally by first infralabials and posteriorly by paired, rectangular postmentals contacting medially for 80 % of their length; one row of enlarged sublabials bordering infralabials, anteriormost largest; gular scales small, rounded, grading into larger, imbricating throat and subimbricate pectoral and ventral scales. Body dorsoventrally depressed, relatively stout (AXG / SVL 0.46); patagium 9.4 mm at midpoint of body, bearing enlarged subimbricate, rectangular scales dorsally, minute juxtaposed, subrectangular scales ventrally, ventral surface bearing raised ridges of granular scales extending from body to edge of flap; 86 minute, flat, round, juxtaposed midbody dorsal scales, largest mid-dorsally; four (L) and one (R) large, flat, isolated, dorsal scale (s) immediately anterior to the hind limb insertions; 42 transverse rows of large, smooth, flat, subimbricate ventrals, ventrals much larger than dorsals, decreasing in size laterally into granular scales at base of flap; 23 enlarged, precloacal scales; five rows of enlarged, post-precloacal scales; and scales immediately anterior to vent granular. Limbs short, robust (FL / SVL 0.11; TBL / SVL 0.15); dorsal scales of forelimbs, flat, juxtaposed, larger than dorsal body scales; ventral forelimb scales subimbricate; anterior and posterior margins of forelimbs, and posterior margins of hind limbs bearing wide, cutaneous flaps; that of anterior margin of forearm (i. e. pre-antebrachial flap) emarginated distally and terminating low on base of digit I; scales of forelimb flap large, elongate, subimbricate; those of hind limb flap much smaller, rounded, subimbricate; palmar scales smooth, rounded; digits fully webbed, relatively short, dorsoventrally compressed; undivided transverse subdigital lamellae number 12 (I), 15 (II), 18 (III), 17 (IV), 18 (V), distalmost lamellae V-shaped; claws arise from within the dorsal surface of digital pads; claw of digit I replaced by an enlarged, flat scale; dorsal scales of hind limbs, flat, juxtaposed, larger than dorsal body scales; ventral scales of hind limbs flat, subimbricate, smaller than ventral scales of belly; flat scales of anterior margin of thigh subimbricate; plantar scales smooth, subimbricate; digits fully webbed; transverse subdigital lamellae number 10 (I), 13 (II), 17 (III), 17 (IV), 14 (V), distalmost lamellae V-shaped; claws arise from within the dorsal surface of digital pads; and claw of digit I replaced by an enlarged, flat scale. Tail original, flattened, shorter than SVL (TL / SVL 0.88); two median rows of transversely widened, smooth subcaudals anteriorly becoming less regular and broken up posteriorly; postcloacal scales large, flat, imbricate; dorsal caudals flat, juxtaposed, larger than dorsal body scales; 6 – 10 smaller scales between much larger, transversely aligned scales composing whorls; tail width and caudal lobes decrease posteriorly; 23 caudal lobes on each side slightly angled posteriorly; and tail terminates in a short, narrow, flap (7.3 mm) bearing weakly crenulated edges. Light phase coloration and pattern in life (Fig. 7). Dorsal ground color of head, body, and tail beige; top of head essentially unicolor; labial scales lighter than body, delimited by thin, dark lines at their junctures; infra-auricular flap same lighter color as labial scales; four faint, thin, sinuous dorsal bands between limb insertions transitioning into approximately six darker caudal bands; terminal caudal band on caudal flap nearly black, edged posteriorly by narrow white band; subcaudal region mottled, weakly banded; iris deep-red; gular region, throat, ventral surfaces of forelimbs, pectoral region, and anterior portion of belly dull-white with stippled scales; and posterior margin of belly dull-white, immaculate; stippling on ventral surfaces of hind limbs dense. Variation (Fig. 8). Variation in coloration and pattern is extensive due to this species’ having very different dark and light phases and its ability to substrate-match (see Grismer 2011 a). Color pattern variation in the paratypes described here is based on preserved material. LSUHC 9059, 9447, and 11418 have more boldly marked dorsal bands. Dorsal caudal bands in LSUHC 11418 are much more prominent than those of the holotype. Dark, subcaudal banding in LSUHC 9447 and 11418 is far more distinct than that of the holotype and in LSUHC 9059 and 10991 the banding is more defined than that of the holotype but not nearly as much as the former three specimens. The belly of LSUHC 11418 is much more densely stippled than other specimens of the type series. The parachute scales of the body patagia do not imbricate in LSUHC 5539, 5597, 6437, 8709, and 9059 whereas in the holotype and the other paratypes they are subimbricate. The tails of LSUHC 5783 and 11058 are regenerated and bear no caudal lobes. Males LSUHC 5597, 5783, 8709, and 11418 have 17 – 25 enlarged pore-bearing precloacal scales. Meristic differences amongst the type material and the additional specimens examined are presented in Table 6. Comparisons (Tables 4, 5; Figs. 3, 5, 6). Ptychozoon cicakterbang sp. nov. differs from all other species of Ptychozoon in having a prominent supra-auricular lobe as opposed to a small ridge or no enlargement at all. It differs further from P. intermedium Taylor, 1915, P. kuhli (Stejneger, 1902), and P. trinotaterra in lacking as opposed to having caudal tubercles. From P. intermedium, P. rhacophorus (Boulenger, 1899), P. trinotaterra, and P. kaengkrachanense it differs in having four body bands as opposed to 0 – 3. Ptychozoon cicakterbang sp. nov. differs from P. bannaense Wang, Wang, & Liu, 2016, P. horsfieldii, P. intermedium, P. kuhli, P. nicobarense, P. rhacophorus, P. trinotaterra, and P. kaengkrachanense in having an emargination between the pre-antebrachial flap and digit I as opposed to no emargination. From P. popaense it differs by having a maximum SVL of 93.4 mm versus 86.2 mm, 14 – 17 subdigital lamellae on the fourth toe versus 13 or 14, the absence versus the presence of a thick, dark postorbital stripe; and the absence versus the presence of large, irregularly shaped, white, vertebral markings. Ptychozoon cicakterbang sp. nov. differs futher from P. lionotum, P. kabkaebin sp. nov., and P. tokehos sp. nov. by having a significantly higher mean number of infralabials and a prominent supra-auricular lobe versus a slightly raised ridge. From C. tokehos sp. nov. it differs further by having a significantly higher mean number of subdigital lamellae on the fourth toe, a higher mean number of ventral scales, and a significantly longer axilla-groin length. From P. lionotum it differs even further by having, as opposed to lacking, prominent ridges on the ventral surface of the patagia and weakly crenulated as opposed to smooth caudal flap edges. From C. kabkaebin sp. nov. and P. lionotum it differs further by having a significantly higher mean number of supralabial scales and midbody scales and caudal whorls composed of enlarged scales. From C. kabkaebin sp. nov. it differs further by lacking, as opposed to having, a thick, dark, postorbital stripe. From C. lionotum it differs further by having a significantly narrower head. Ptychozoon cicakterbang sp. nov. is well-separated from other species previously recognized as P. lionotum in the PCA and DAPC and occupies a significantly different position along PC 1 from that of P. kabkaebin sp. nov. and P. tokehos sp. nov. and along PC 2, it occupies a significantly different positon from that of P. lionotum. From these three species, it is further separated by an uncorrected pairwise sequence divergence of 4.1 – 14.4 %. Combinations of other characters differentiating P. cicakterbang sp. nov. from other more distantly related species are presented in Table 5.
Grismer, L. Lee, Wood Jr, Perry L., Grismer, Jesse L., Quah, Evan S. H., Thy, Neang, Phimmachak, Somphouthone, Sivongxay, Niane, Seateun, Sengvilay, Stuart, Bryan L., Siler, Cameron B., Mulcahy, Daniel G., Anamza, Tashitso, Brown, Rafe M. (2019): Geographic structure of genetic variation in the Parachute Gecko Ptychozoon lionotum Annandale, 1905 across Indochina and Sundaland with descriptions of three new species. Zootaxa 4638 (2): 151-198, DOI: 10.11646/zootaxa.4638.2.1