Adeonellopsis japonica
(Ortmann, 1890)
GBIF:164249544
0
Synonyms
ABOUT
Descriptions(5)
Export occurrence data
Darwin Core Archive (ZIP)
CLASSIFICATION
Taxonomic Classification Tree

MULTIMEDIA
Media Files(4)
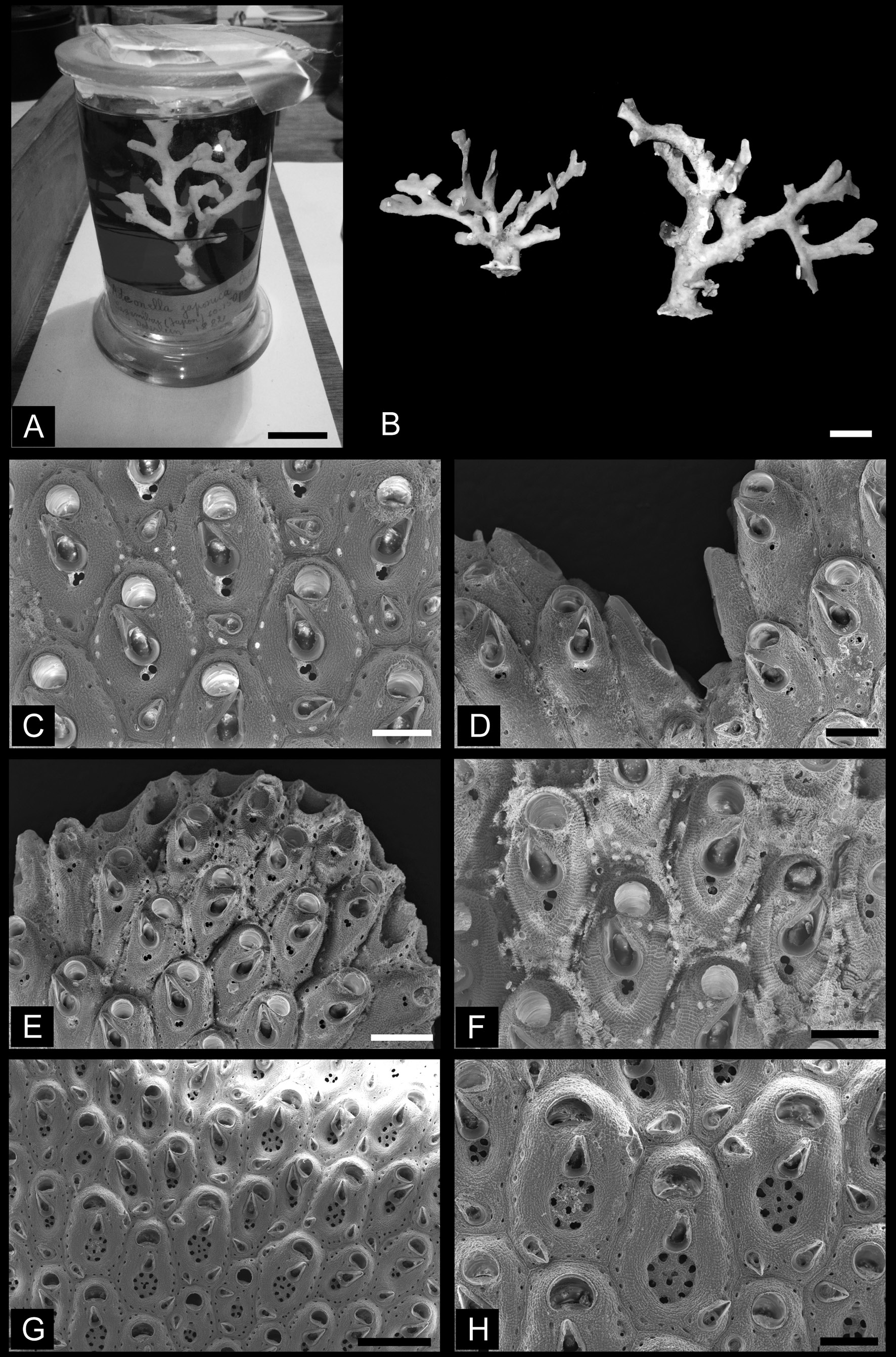
Fig. 7. Adeonellopsis japonica (Ortmann, 1890). A. Lectotype from among Ortmann’s specimens in the Döderlein Collection (MZS 3-3). B. Colonies collected at Otsuchi (NSMT-Te760). C. Autozooids with large suboral avicularium and biporous spiramen (NSMT-Te741, from lectotype, MZS 3-3). D. Lateral vicarious avicularia at branch bifurcation (NSMT-Te741, from lectotype, MZS 3-3). E. Young autozooids at distal end of branch (NSMT-Te741, from lectotype, MZS 3-3). F. Young autozooids, with biporous spiramen surrounded by thick, swollen rim bearing rows of granulation (NSMT-Te741, from lectotype, MZS 3-3). G. Autozooids and gonozooids at branch bifurcation (NSMT-Te742, from paralectotype MZS 3-4). H. Gonozooids showing large multiporous spiramen (NSMT-Te742, from paralectotype MZS 3-4). A–B = optical photographs; C–H = SEM images. Scale bars: A = 2 cm; B = 1 cm; C–D, F, H = 200 μm; E = 300 μm; G = 500 μm.

Fig. 8. Adeonellopsis japonica (Ortmann, 1890). A. Distal end of branch just starting to bifurcate, showing young autozooids and gonozooids (NSMT-BryR277). B. Enlargement of young gonozooids, showing peripheral rim with rows of granulation perpendicular to margin, and large multiporous spiramen (NSMT-BryR277). C. Autozooids in middle of branch, showing immersed spiramen (NSMT- Bry R 193). D. Basal part of branch, showing autozooids, and some kenozooids with closed orifice (NSMT-BryR36). E. Young gonozooids, showing distinct marginal pores and multiporous spiramen (NSMT-Te758). F. Distal end of branch, showing autozooid formation (NSMT-Te758). SEM images. Scale bars: A, C = 1 mm, B, D–E = 300 μm; F = 500 μm.

Fig. 9. Adeonellopsis japonica (Ortmann, 1890). A. Colony collected at Albatross Station D, off Oahu Island, Hawaii (USNM 271601). B. Autozooids of Hawaii specimen, showing suboral avicularia and biporous spiramen (USNM 271601, Hawaii). C. Autozooids and lateral vicarious avicularia at a branch bifurcation (USNM 271601 paper box, Hawaii). D. Enlargement of lateral vicarious avicularia at branch bifurcation (USNM 271601 paper box, Hawaii). A = optical photograph; B–D = photomicrographs. Scale bars: A = 5 mm; B, D = 300 μm; C = 500 μm.

IMAGES