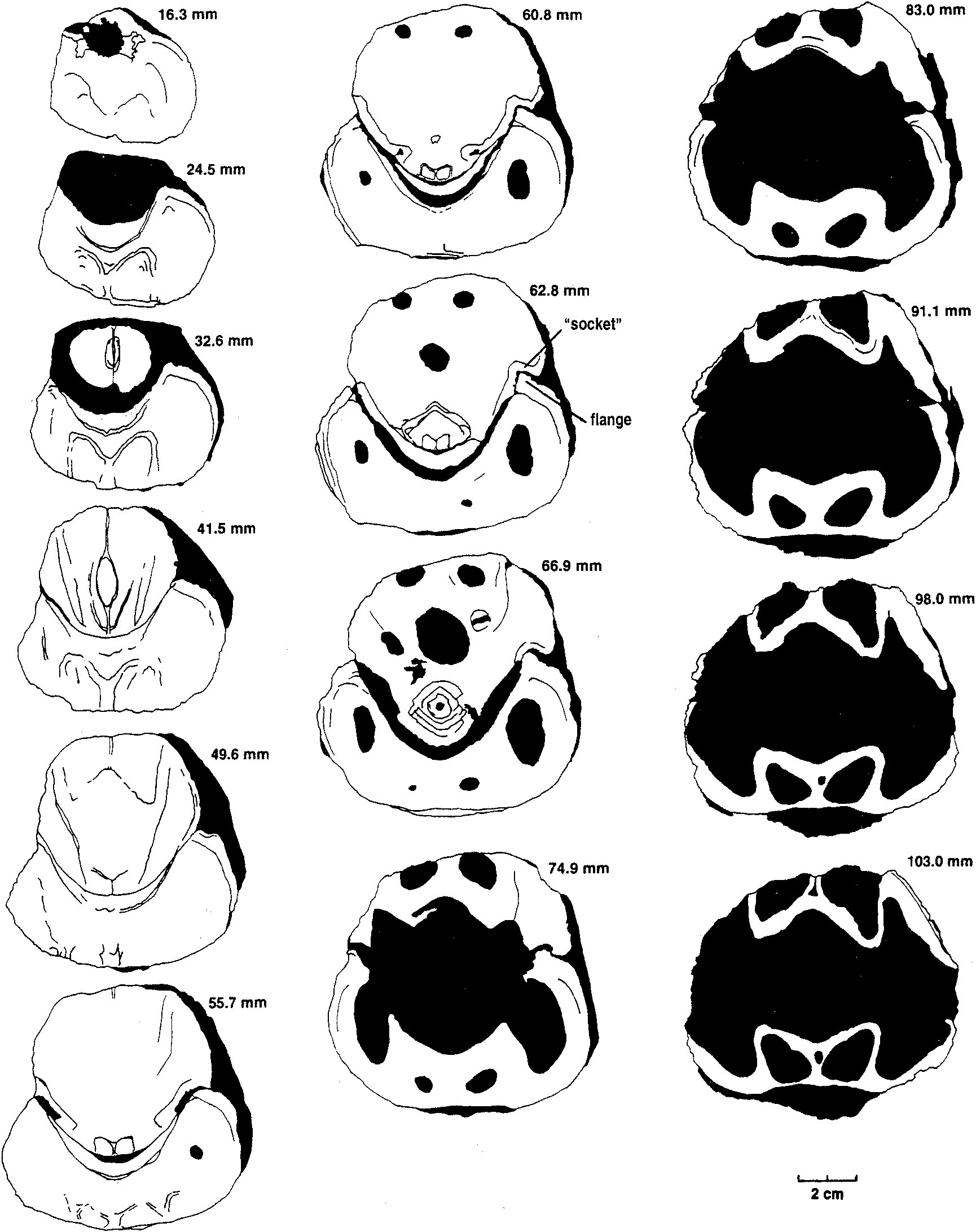
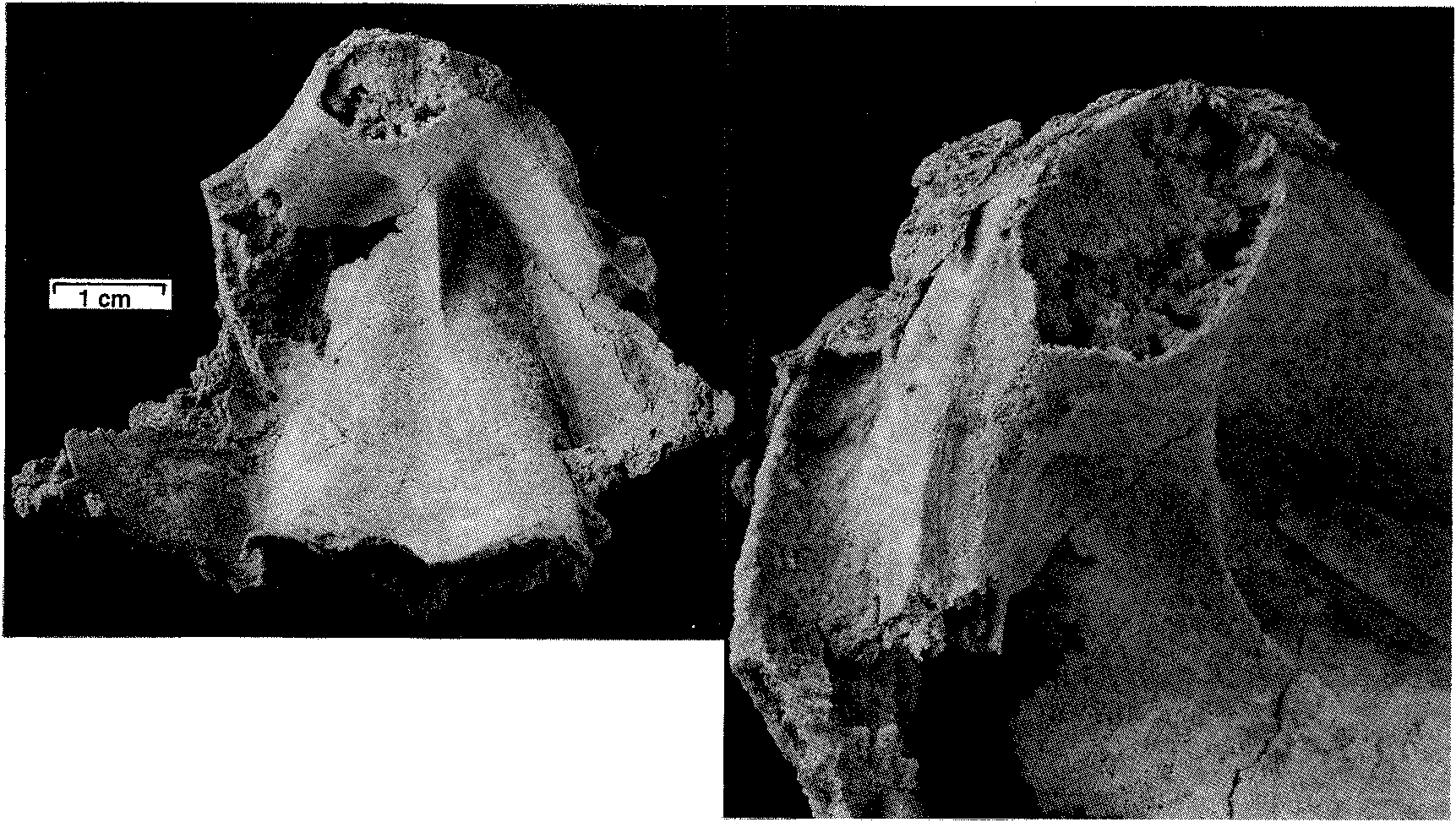
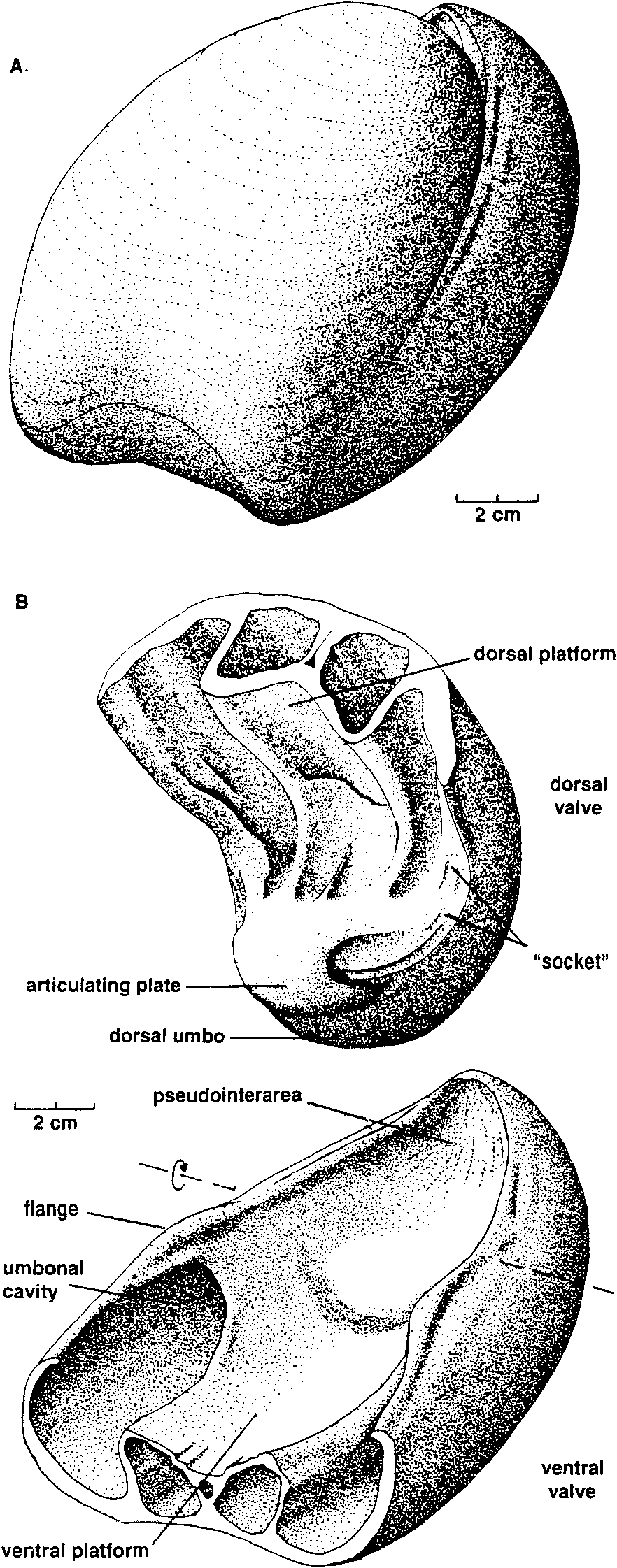
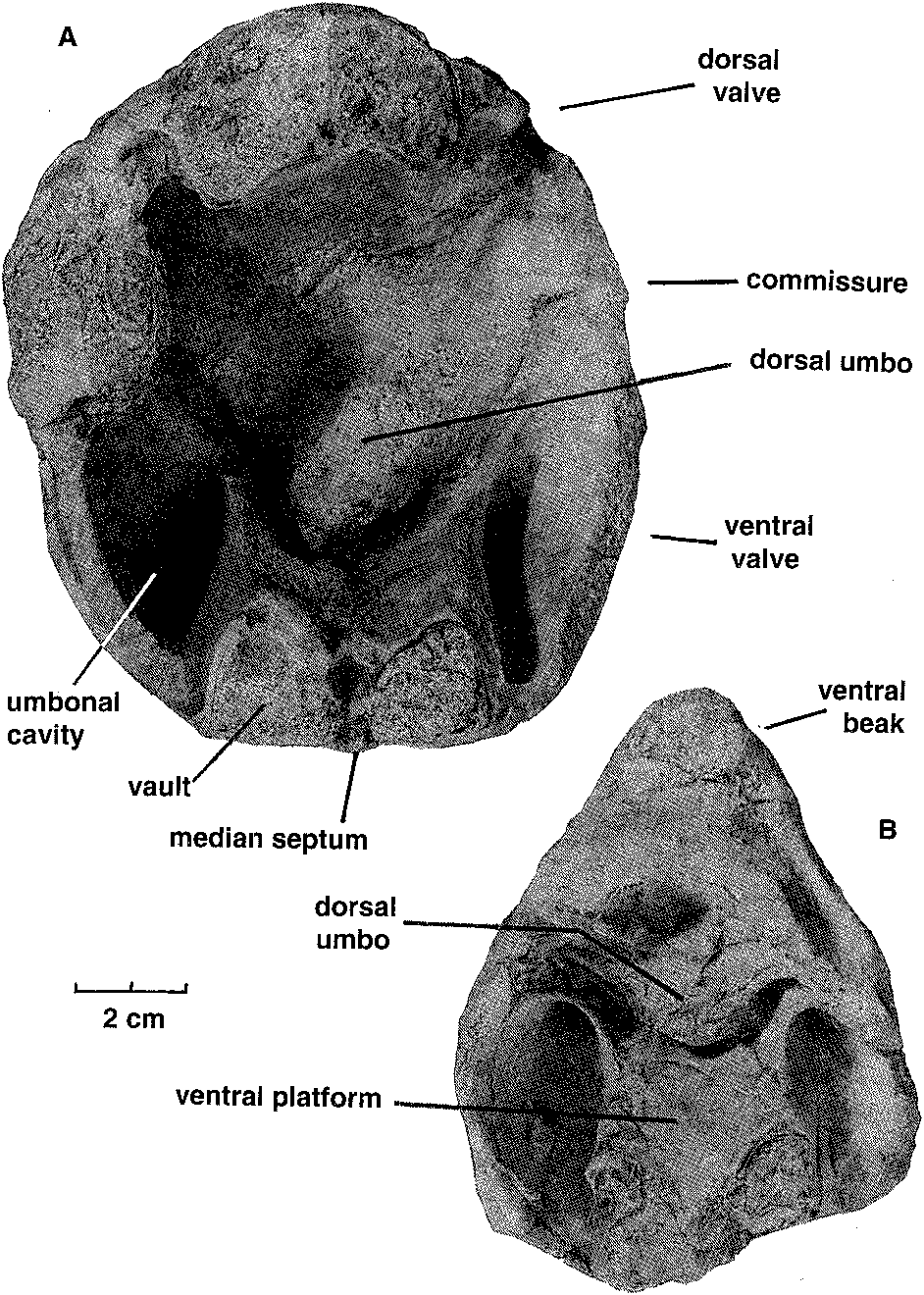
mm), a low flange develops on the inner edge of each ventral valve margin which fits into a groove near the inner edge of each dorsal valve margin. The inner edges of the grooves disappearforwards, leaving the tapered outer edges of the dorsal valve margins resting against the outer surfaces of the ventral flanges. The flanges in turn become lower, the ventral valve margins narrow, and from about the shell mid-length the commissure assumes a more conventional appearance, without any overlap of dorsal valve over ventral valve. Ventral interior: The ventral platform is strongly developed, about 40 % as high as the valve and occupying about a third of the total width anteriorly. It extends well beyond mid-length, and covers a large cavity which is divided into two long vaults by an even longer median septum which may reach the start of the anteroventral sulcus. In cross-section the sides are steep to slightly overhanging, and gently convex, while the upper surface is a broad shallow trough which longitudinal sections and the excavated shell show is not a continuous smooth curve in front of the contact zone with the dorsal umbo, but terraced, the faces of the terraces facing forward. The one silicified ventral valve fragment (MMF 33367) shows a strong but fairly smooth bulge in a position just forward of where the remnant of the dorsal umbo rests against the pseudointerareaPosteriorly, the upper edges of the platform rise steeply and flare outwards then forwards to merge with the valve margins in continuity with (and so buttressing) the marginal flanges which interlock with the dorsal valve. They thus enclose deep umbonal cavities whose apices are about level with or slightly behind the flattened pivot surfaces on the valve margins (Fig. 6). The platform vaults are increasingly constricted posteriorly by thickening of the top and sides of the platform; from the sections, the vaults start in front of the umbonal cavities. There is also a narrow and relatively short conical cavity at the junction of the platform and median septum. No muscle impressions have been seen on the available specimens. Dorsal interior: The dorsal platform is similar in extent and form to that in the ventral valve, including a strong median septum and long vaults, but its edges merge posteriorly with the swollen beak rather than with the valve margins. The platform surface appears to be smooth, without any indication of muscle insertion. Umbonal cavities are apparently absent but, as with the ventral platform, there is a narrow cavity at the front of the platform, at its junction with the median septum. There is a close fit between the dorsal umbo and the posteriorly raised margins of the pseudointerarea (Figs. 5,6). Partly enveloping the dorsal beak is a thick curved plate (see Fig. 8), which arises from a large mass of dense tissue resting on the posterior end of the dorsal platform. This plate is concentric with, but raised above, the dorsal umbo, and extends longitudinally through an arc of about 120 to 1500, such that it approaches the surface of the ventral platform. In paratype MMF 33366 its base is supported by a distinct ridge extending straight up from the surface of the platform; this, presumably the cardinal buttress, is not evident in the transverse sections. Silicified paratypes MMF 33365 and 33366 (Figs. 9, 10) show that the laterally placed recesses between plate and umbo are striated; they are presumed to have been sites of diductor muscle attachment (see discussion below). Longitudinal sections (Fig. 11) show that the distal end of the articulating plate faces a terrace on the ventral platform. Beneath that terrace distinct closely spaced growth traces can be seen, and the tissue here is clearly a pad resting on the surface of the ventral platform, rather than being part of the platform structure itself; it probably corresponds to the swelling seen in MMF 33367. Dimensions (Table 1). There are only one or two more or less complete shells, all others being damaged. In most cases only the posterior half (more or less) of the shell is preserved, and examination of the outcrop reveals that this is a function of the living position. The shells occur in relatively crowded layers, mostly beak downwards, and each layer has been truncated by storm action, removing or damaging the anterior ends of the larger shells. In many shells the valves are gaped and slightly dislocated. The originally probably aragonitic shells are recrystallised (at times very coarsely-see Figs. 8, 11) to calcite, and weathered-free shells have generally lost part ormore often all of the outer surface. Consequently in most cases the dimensions for width and depth shown in Table 1 are approximate, and for length or depth an estimate using the one complete shell provides a guide to probable outline. Discussion. Few described trimerellides approach Keteiodoros bellense n. sp. in size or convexity. Closest are the gigantic Belubula Percival, 1995, from the Upper Ordovician Belubula Limestone southwest of Orange, NSW, and the somewhat smaller Adensu Popov & Rukarishnikova, 1986, from rocks of similar age in southern Kazakhstan. Belubula, estimated to have reached a length of 20 cm, is also comparable with Keteiodoros in its deep ventral umbonal cavities, but the ventral platform is low, not vaulted or excavated, and there is a stout cardinal buttress; the dorsal platform, too, is relatively low. At a length of about 90 mm and a depth of 50 to 70 mm, Adensu is only half the maximum size of the Australian shells. The genus was made the type of a new trimerellide family based partly on large size and great convexity, but mainly in having raised dorsal muscle fields separated by deep grooves, and lacking the typical trimerellide platforms in both valves. Percival (1995) did not think the available material justified a separate family, and pointed out similarities with gerontic Eodinobolus stevensi Percival, 1995, from the Fossil Hill Limestone. While Trimerella and Dinobolus have vaulted platforms in both valves, their shells are much less convex, the ventral beaks are high and not strongly incurved, with well differentiated pseudointerareas, and the dorsal umbos are small. In all these features they are unlike Keteiodoros. Articulation in Trimerella is obscure but, in Dinobolus, Eodinobolus and Gasconsia, and possibly Monomerella and Adensu, it appears to comprise a transverse cardinal socket overhanging the platform in the ventral valve, in whichrests the posterior edge of an articulating plate which is effectively the thickened and protruding posteriormargin of the dorsal valve (Norford, 1960; Norford & Steele, 1969; Hanken & Harper, 1985; Popov & Rukavishnikova, 1986; precise details of the articulation have been questioned by Mergl, 1989). The simpler term " hinge plate " is already used for part of the cardinalia in rhynchonellate brachiopods. The arrangement in Keteiodoros is seemingly more complex: it uses areas on the valve margins as pivoting surfaces, while medially there is articulation between the ventral surface of the dorsal umbo on the one hand, and the surface of the pseudointerarea on the other, a little like a unidirectional " ball and socket " joint; the articulating plate appears to have served mainly as a " stop " to unwanted longitudinal movement, and as a protective cover for the diductor muscle attachment. Possibly related is Monomerella ovata Whiteaves, 1884, from the Ludlow-age Guelph Dolomite of Ontario (Holmer, pers. comm., 1997). Dorsal internal structures, critical to generic assignment, have not been described, but the ventral interior illustrated by Hall & Clarke (1892, pI. IVD, fig. 15) differs in its long well-differentiated pseudointerarea and much shorter platform. In Australia, Talent (pers. comm., 1995) has collected rather smaller but externally very similar specimens from the Wenlock Ludlow Yarrangobilly Limestone in New South Wales, and the Silurian at Chillagoe in northern Queensland; these remain undescribed. Finally, it is interesting to compare the quite specialised Keteiodoros, with its complex morphology and gigantic size, with Ussunia Nikitin & Popov, 1984, from the late Llandeilo to early Caradoc of Kazakhstan. An early trimerellide-like genus displaying features transitional from craniopsids, Ussunia is biconvex but not at all globose, with an undifferentiated pseudointerarea extended anteriorly as a flattened limbus. Muscles were inserted directly on to the valve floor-platforms were not developed. The shell was thick and solid, without umbonal excavation to lighten the valves. Ussunia has no form of mechanical articulation, and the dorsal beak showed no sign of enrolling. Keteiodoros averages 4 to 5 times the length and width of Ussunia, and is about 7 times as globose. The two genera are separated in time by about 35 million years; Ussunia represents the small unspecialised ancestral stock, while Keteiodoros is the highly specialised, complex and gigantic descendant on a line soon to pass into oblivion. Cope's Rule was rarely more clearly expressed. Functional morphology. The most distinctive feature of this new trimerellide, apart from its size and strong biconvexity, is the inferred method of articulating the valves. This involves the valve margins, the dorsal umbo, and the pseudointerarea. Polished and serial sections (e. g., Figs. 6, 11), and prepared shells (Fig. 7), show that the dense posterior part of the dorsal valve, greatly enlarged and extremely strongly incurved, fitted laterally within the limits of the posteriorly equally dense ventral platform and pseudointerarea. Infront of this region where the dorsal umbo sat within the pseudointerarea over its full width, the lateral margins of the dorsal valve expanded rapidly, such that a solid shelf formed on each side which rested on the flattened top of the ventral valve margin (Fig. 6, from 55.7 mm; Fig. 8). This zone of flattened valve margins was short, and was where the margins crossed over one another as seen dorsoventrally. These flattened zones are inferred to have acted as pivots (not teeth and sockets) for the opening and closing of the shell. This is a form of articulation which, while rudimentary, is nevertheless somewhat reminiscent of that in some non-strophic rhynchonellate shells such as pentamerides. In front of the pivot zone each dorsal valve margin developed a broad somewhat oblique groove, into which fitted a similarly oblique flange arising from the inner edge of the ventral valve margin (Figs. 6, 7, 12). Functionally, the result was an interlocking system which, when the shell was closed, prevented relative movement of the two valves in the plane of commissure. When the shell was open, the close fit of the dorsal umbo in the pseudointerarea probably also constrained by the position of the adjacent muscles, would have provided significant restraint on relative lateral and skewing movements between the valves. None of the available material is well enough preserved to show muscle scars but, if one considers the musculature involved in shell articulation, it can be assumed that the adductors would have extended from one platform to the other, as inferred for other trimerellides (Norford & Steele, 1969, text-fig. 2; Mergl, 1989). Given the size and probable weight of the valves, the considerable strength of such short thick muscles would probably have been very necessary to close the shell, and even more so to keep it closed against adverse conditions. However, the method of opening the shell poses problems, especially in view of the close fit between pseudointerarea and dorsal umbo. Muscles extending forward from the anterior surface of the dorsal umbo to the ventral platform upon contraction would have served not to open the shell, but simply to pull the dorsal valve forward (which the interlocking of the valve margins and the bulge on the ventral platform in front of the dorsal umbo would have prevented anyway). To achieve proper leverage, muscles operating as diductors (we have no way of establishing precise homologies, so in this paper use the term " diductors " in a purely functional sense, as the muscles responsible for opening the shell) must have extended forward from somewhere on the dorsal umbo behind or ventral to the above-described marginal pivot zones. Muscles attached to the distal face of the curved articulating plate and extending to an attachment area well forward on the ventral platform would have met this criterion, but there are two difficulties: when the shell was closed, there would have been little if any room between plate and platform for passage of the muscles, and contraction of the muscles would have put significant strain on the base of the plate, possibly enough to break it. Examination of serial and longitudinal sections, and of the silicified specimens, suggests an alternative (Fig. 13): that the dorsal ends of the diductors were attached to the umbonal surface in the lateral slots at the sides of the articulating plate, and emerged to either side of the plate. There are appropriate gaps at the sides of the dorsal umbo between sections at 55.7 and 60.8 mm in Fig. 6, approximately aligned with, but ventral to, the marginal pivot zones, and also in the peels from MMF 33364, Fig. 8. As already noted, MMF 33365 and 33366 reveal appropriately situated striated areas suitable for such attachment. With the diductors attached to the outer parts of the ventral platform in front of its mid-length, there would have been adequate space for the viscera and adductors medially. Mechanically this would have formed an effective lever system, opening the shell about the proposed pivot points to at least a 10 ° gape and possibly as much as 20 °. However, one effect of such a system is that the dorsal umbo would have slid on the pseudointerarea unless restrained by soft tissue and periostracum. The geometry visible in longitudinal sections would have allowed for this movement, and the posterior geometry of the shell appears to be such that neither the exterior space between the two umbos (which is in any case not very large) nor the visceral space posterior to the dorsal umbo changed significantly during such a movement. The laminated pad on the floor of the pseudointerarea occupying the zone where such sliding would have occurred, may have formed a bearing surface for such movement, perhaps also serving by a close fit of the two surfaces to exclude sediment. Moreover, the articulating plate would have protected the dorsal ends of the laterally placed muscles from damage during such movement. A thickened zone of periostracum may have been present, but this is uncertain because of the state of preservation of the shells. The manner of posterior closure, and the way in which the valves moved during opening, is important, because the shells sat freely umbo-down in sediment, and the space between dorsal and ventral umbos could only have been occupied by either tissue or silt. There appears to be no room forpassage of a pedicle (in common with othertrimcrcllidcs), and so it is unlikely there was organic tissue filling the space, but unless the silt were either extremely soft and fluid (thixotropic?) or shaped bythe animalinto afirmstablecavity, it would have been an impediment to valve opening. The proposed opening mechanism would not have been strong enough to work against loose but resistant material, and the diductors would probably have served (in conjunction withrelaxation of the adductors) simply to initiate opening. This is supported by the observation that the weight distribution of the dorsal valve about the pivots appears to have been evenly balanced when the shell was closed. The large solid beak on the ventral side of the pivot-line was balanced by the larger volume of thinner-walled platform and shell wall making up the anterior part of the valve. Stability in that position was increased by the solid shell mass making up that part of the dorsal valve posterior to (hence below) the pivot-line. However, once opening movement was initiated, the weight of the anterior part of the dorsal valve, moving through a perceptible arc, would have overcome the weight of the dorsal umbo which barely changed its orientation relative to the pivot zone. The considerable weight of the dorsal valve would have required the constraint of strong adductor muscles to stop it from sliding downwards out of the bearing surface formed by the pseudointerarea. The size and elevation of the platforms suggests that the adductors were indeed strong, but this function may also have been aided by the close conjunction of the distal end of the articulating plate with the terraced face of the tissue pad which formed the bearing surface on the top of the pseudointerarea. Moreover, at least in gerontic shells the strongly incurved ventral beak formed a cup effectively restraining the dorsal valve from below. A further implication of the weight of the valve is that closing the shell would at least initially have required the strong adductors suggested above. Phylogenetic relationships The new genus is characterised by gigantic size and a unique articulatory mechanism comprising dorsal articulating plate, robust zones on the edges of the ventral valve pivoting against equally robust zones on the dorsal margins, and, in front of those zones on the valve margins, oblique ventral flanges restraining twisting movement by articulating with marginal dorsal " sockets " (Fig. 12). These and other associated morphological features adapted Keteiodoros well to its preferred environment. Taken by themselves, tendencies towards gigantism or oblate / spheroidal shape have relatively little phylogenetic significance. For example, Adensu Popov & Rukavishnikova, 1986, is a monotypic Late Ordovician trimerellide, endemic to Kazakhstan, which has a similar shape to, and dimensions approaching those of Keteiodoros. However, Adensu lacks both raised muscle platforms and umbonal cavities, and its articulating plate lies parallel to the commissure. Accordingly it is not thought to be closely related to Keteiodoros. Substantial umbonal shell thickening is a typical means in many unrelated brachiopods (e. g., pentamerides) of preventing dislodgement by waves or currents of shells lacking pedicle or cementation attachment. Long umbonal cavities are likewise common to most known Silurian trimerellide genera, as are prominent ventral platform vaults. The only known trimerellides with strongly vaulted dorsal platforms are Paradinobolus Li & Han, 1980 and Prosoponella Li, 1984 (bothfrom the early Ashgill of South China), and the widely distributed and long-ranging Trimerella (late Ordovician to middle Silurian). A single silicified incomplete dorsal valve with such a platform, referred doubtfully to Paradinobolus, is also known from the Late Ordovician Malongulli Formation in central New South Wales. Superficial similarities aside, none of these trimerellides share sufficient characters in common with Keteiodoros to be considered directly related. The specialised articulatory mechanism of Keteiodoros is likely to provide the best clue to its evolutionary relationships. One trimerellide, Belubula spectacula Percival, 1995, from the Late Ordovician (early Eastonian, equivalent to late Caradoc) Belubula Limestone of central New South Wales, is a potential ancestor to Keteiodoros. Although separated in time by approximately 25 million years, and without intervening evolutionary stages being known at present, the two genera share significant morphological similarities, habitat preferences, and geographic proximity. Belubula is of gigantic size for Late Ordovician trimerellides, reaching an estimated 200 mm in length and 100 mm in width; the largest individuals are thus comparable in length with Keteiodoros, though the latter is relatively much wider and deeper. In profile, both are similarly strongly equibiconvex. Note also that both genera display asymmetry (compare Percival, 1995: Fig. 4 J with MMF 33365, Fig. 9), although the direction of asymmetry of only the ventral valve of Belubula is apparent from the figure. Grossly thickened posterior regions of the ventral valves in both Keteiodoros and Belubula contain large conical umbonal cavities. Of particular significance is one gerontic individual of Belubula (Percival, 1995: Fig. 4 C) which exhibits an incipient modification of the dorsal homeochilidial (or articulating?) plate which could well have led to the specialised articulation of Keteiodoros. Percival (p. 50) noted in reference to this particular silicified dorsal valve that in gerontic specimens this plate " can enlarge to form a prominent curved bulbous plate, bearing growth striations, & extending ventrally above level of commissure with a knife-edge anteroventrally ". It is reasonable to suggest that over the ensuing 25 million years of evolution this plate developed into the robust incurved articulating plate of Keteiodoros, bearing a median bulge and buttressed anteriorly by a median ridge, and forming part of a large ball-like dorsal umbo bearing against the concave pseudointerarea. Whereas a massive muscle (attached to the ventral cardinal socket and inserted in the dorsal subcardinal depression) was essential in Belubula to prevent a shearing motion of the valves during articulation, paired diductors attached at the sides of the articulating plate, lateral to the ball joint, could have served the same purpose in Keteiodoros. ACKNOWLEDGMENTS. In reconciling the initially divergent ideas on aspects of this unusual brachiopod, a large part has been played by the technical help we have received, and for which we are grateful. Michael Doyle of AGSO had the difficult task of mechanically preparing specimens, made the first set of serial sections, and did much of the photography. At the University of Wollongong Andrew Southall prepared peels from the serial sections, and Trudi Marshall prepared drawings from those peels. Gary Dargan of the Geological Survey ofN. S. W. prepared further sections and the silicified specimens, while David Barnes photographed them. We would also like to thank Lars Holmer and reviewers Michael Bassett and Jisuo Jin for their interesting and useful comments. John Pickett and Ian Percival publish with permission of the Director-General, New South Wales Department of Mineral Resources. Desmond Strusz completed the paper while a Visiting Fellow in the Geology Department, Australian National University, and would like to thank Prof. Arculus for providing the necessary facilities.
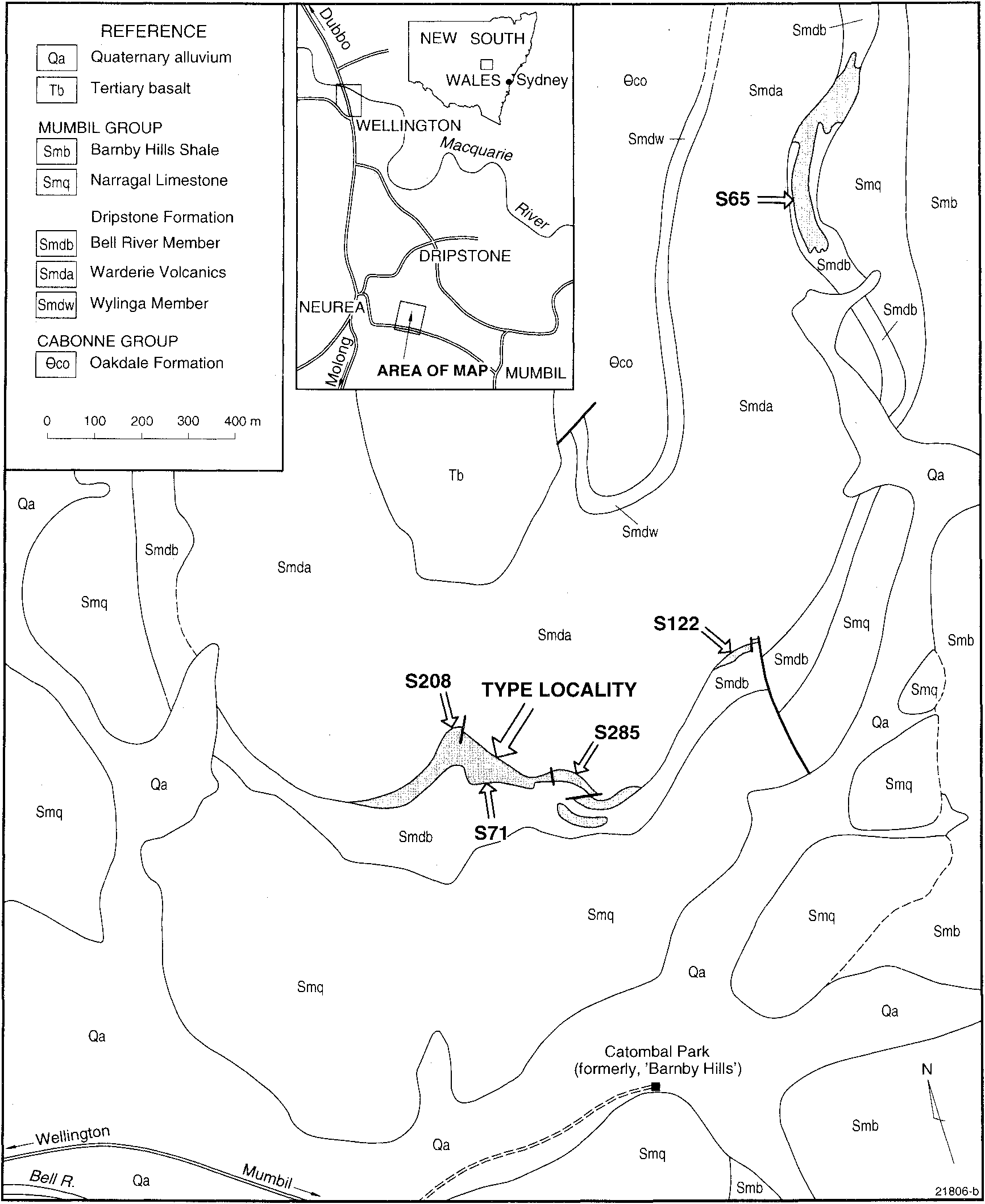
Strusz, D. L., Percival, Ian G., Wright, A. J., Pickett, John W., Byrnes, A. (1998): A giant new trimerellide brachiopod from the Wenlock (Early Silurian) of New South Wales, Australia. Records of the Australian Museum 50 (2): 171-186, DOI: 10.3853/j.0067-1975.50.1998.1278, URL: https://journals.australian.museum/strusz-et-al-1998-rec-aust-mus-502-171186/