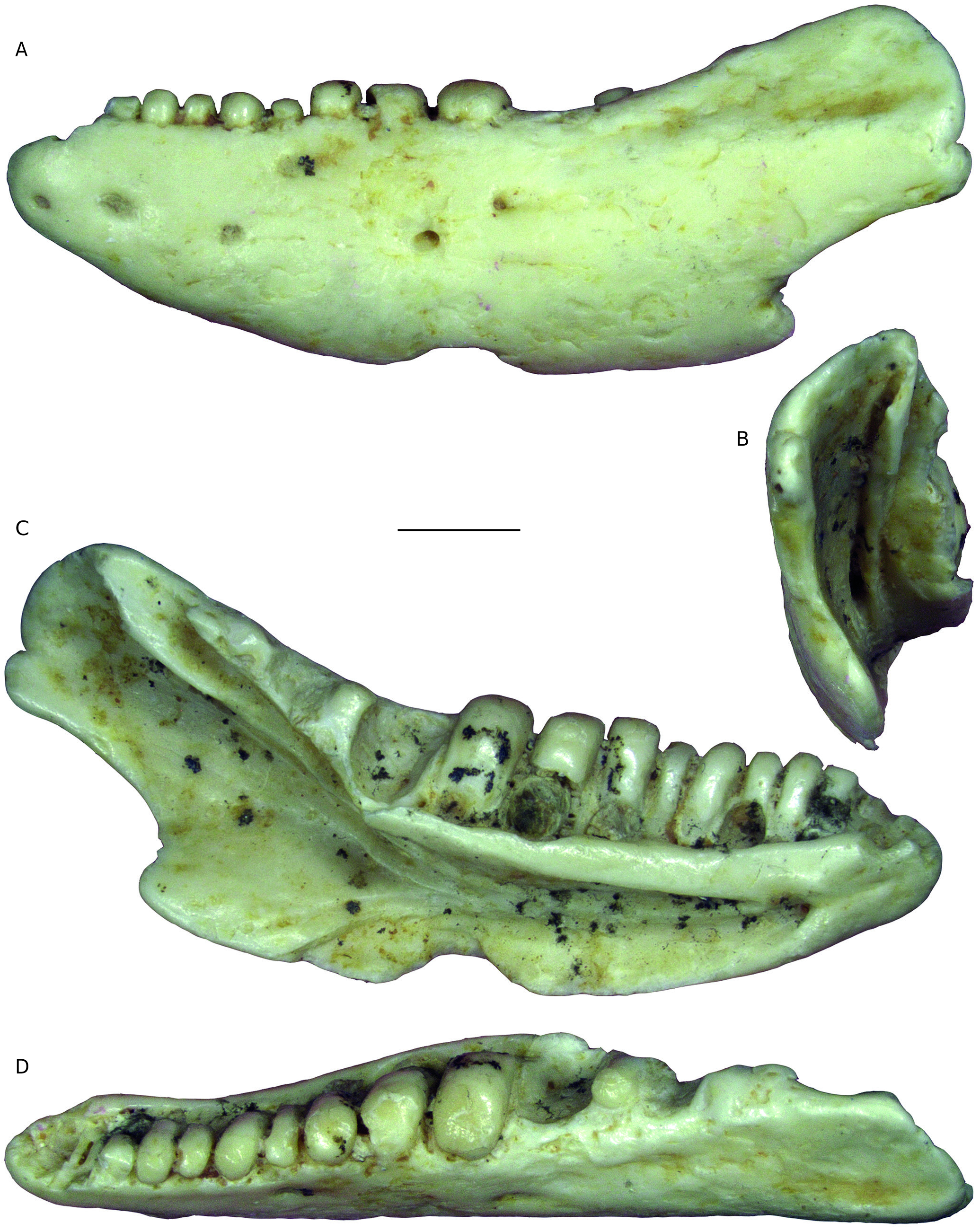
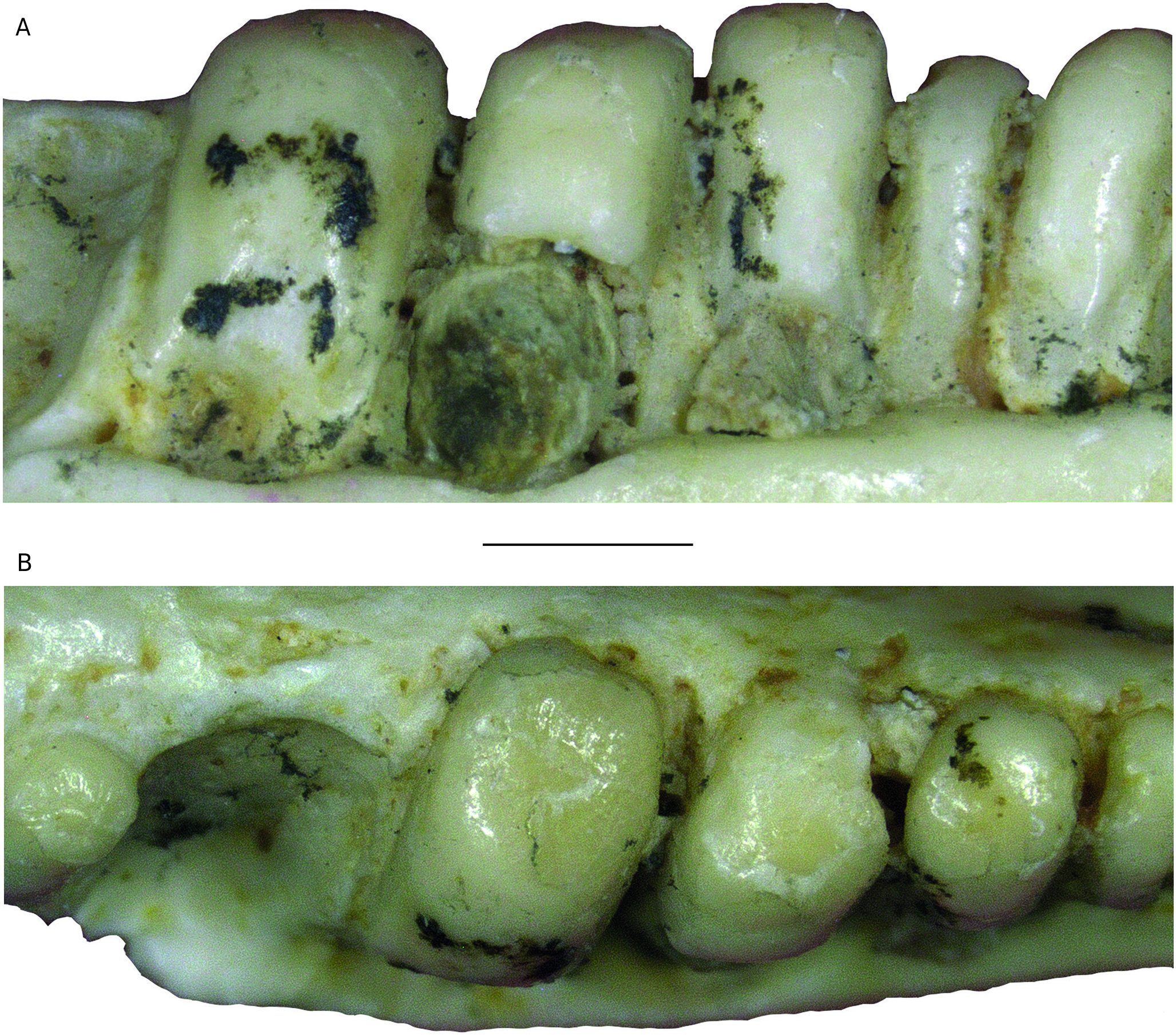
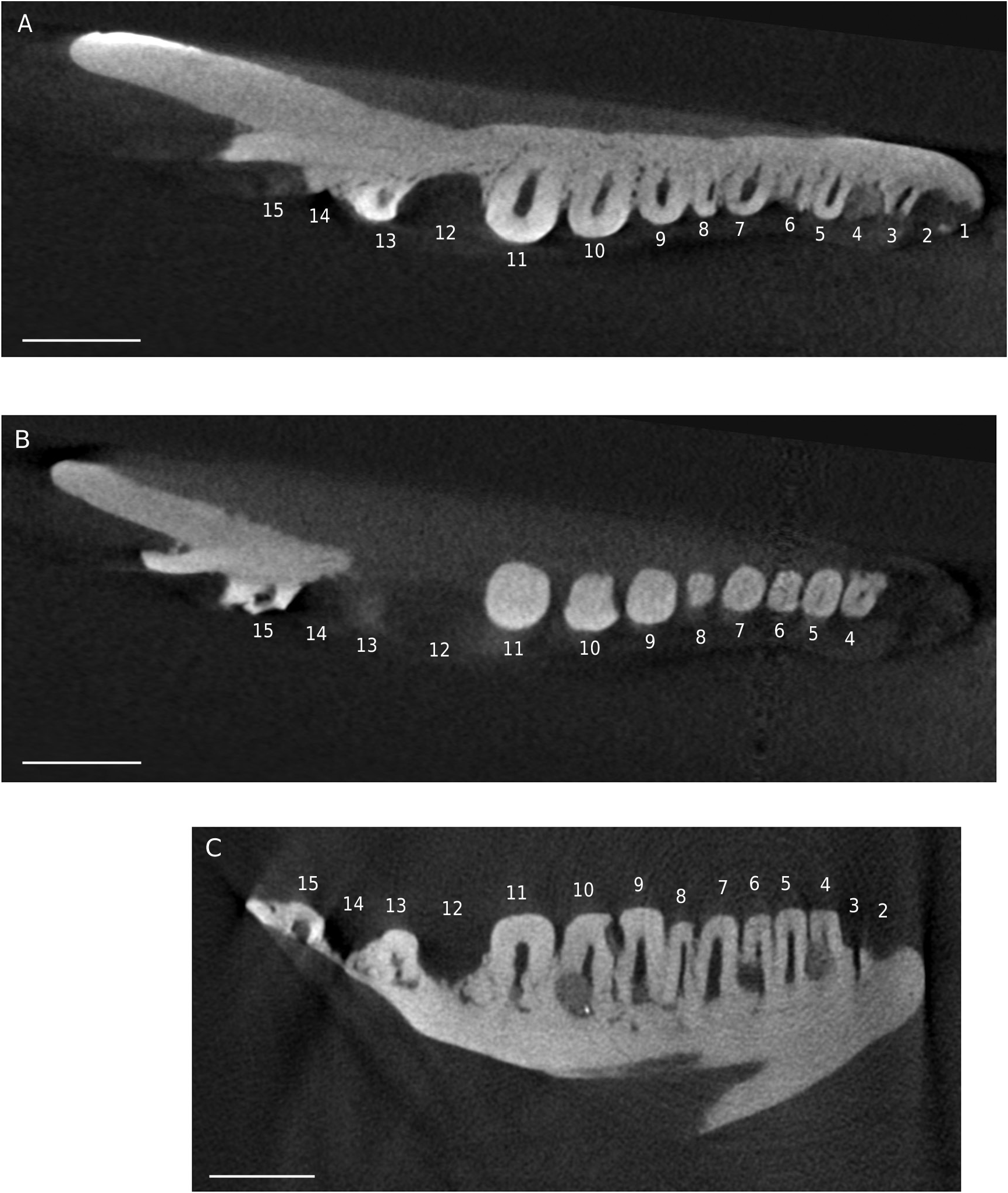
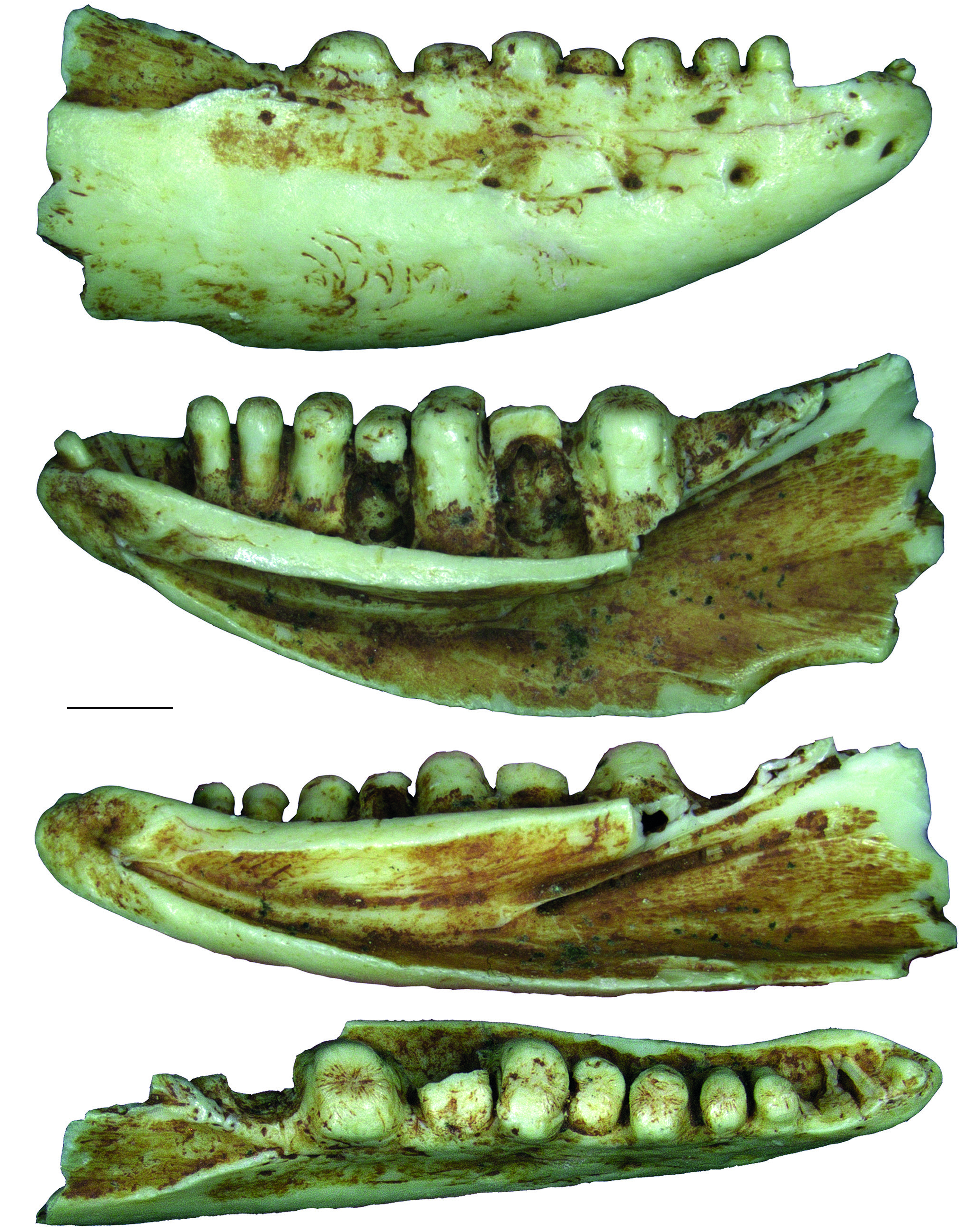
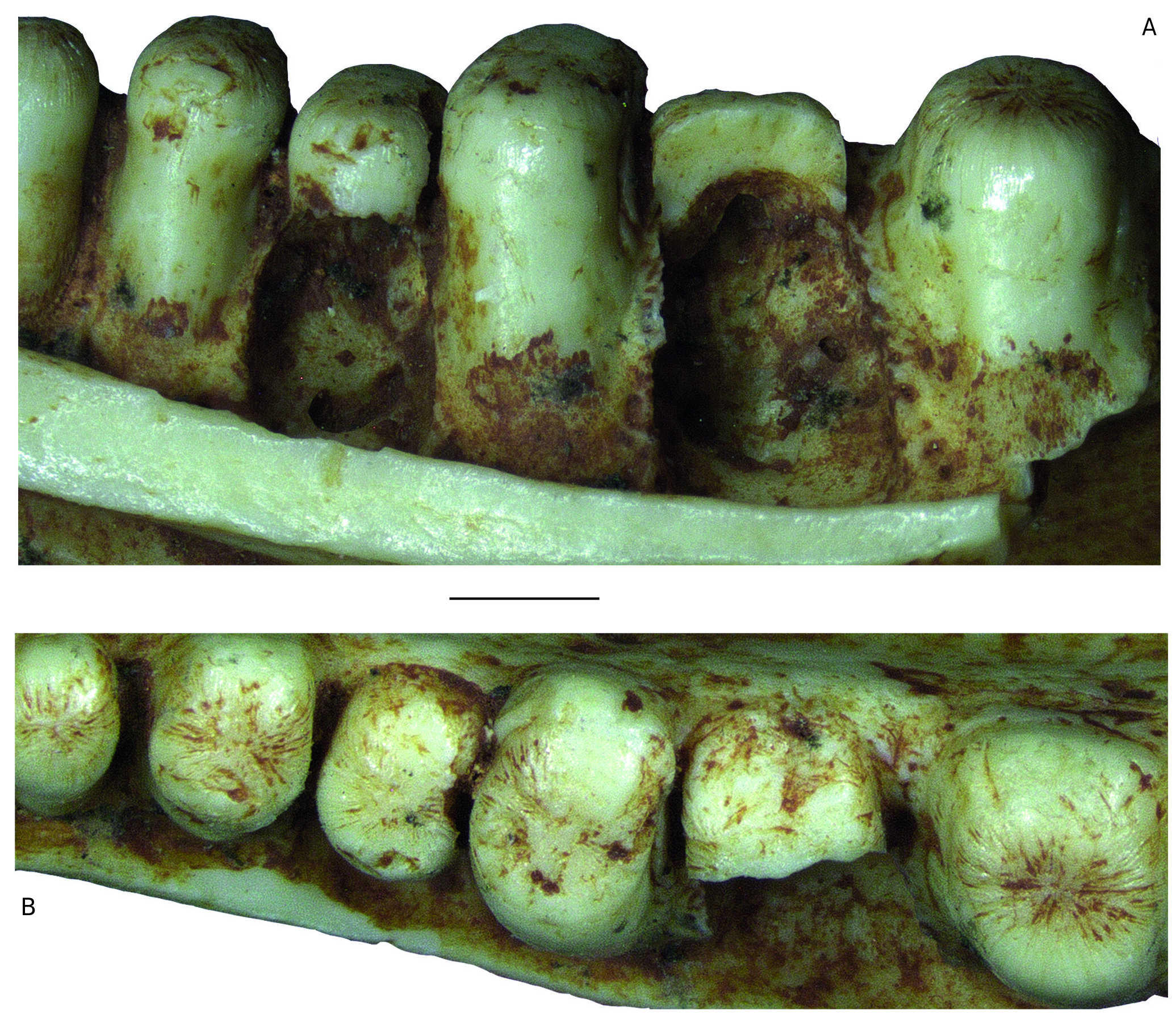
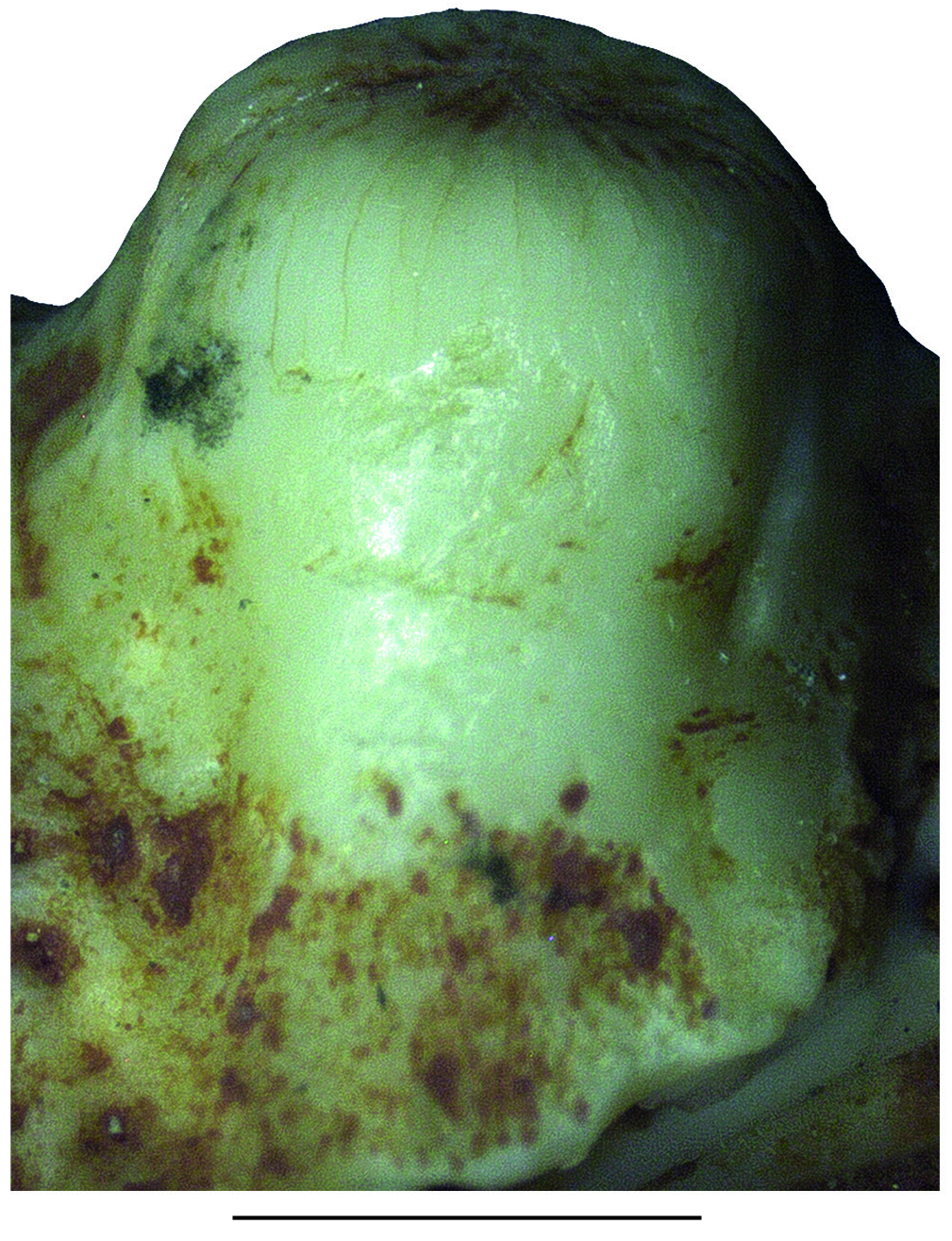
REMARKS Pseudeumeces cadurcensis was originally established by Filhol (1877 a) as a scincid of the extant genus Plestiodon Duméril & Bibron, 1839. This taxonomic opinion was subsequently followed by others (e. g., Nopcsa 1908; Kuhn 1939), although affinities with the glyptosaurine Placosaurus also appeared in the literature (Lydekker 1888 b; Leenhardt 1926). Hoffstetter (1944) was the first to realize its lacertid affinities and placed it into its own genus, Pseudeumeces. So far, Pseudeumeces cadurcensis represented the only currently recognized species of this genus, as other two species that have in the past been referred to Pseudeumeces are now known to pertain to other genera or represent indeterminate lizards (i. e., Glyptosaurus walbeckensis Kuhn, 1940, which was recombined into Pseudeumeces by Estes [1983], and Pseudeumeces pouiti Augé, 1993, which was subsequently recombined to its own genus Ligerosaurus Augé, Bailon & Malfay, 2003, as Ligerosaurus pouiti by Augé et al. [2003]). We consider that the dentition and overall morphology of our new species Pseudeumeces kyrillomethodicus n. sp. appears to bear a resemblance with Pseudeumeces cadurcensis. Similarly to the case of our new taxon, the type material of Pseudeumeces cadurcensis also originates from an imprecise locality within the Phosphorites du Quercy (Filhol 1877 a; for this taxon see also Augé 2005; Augé & Hervet 2009; Čerňanský & Augé 2012; Čerňanský et al. 2016 a; Bolet et al. 2017). Nevertheless, the material described herein clearly exhibits several obvious differences relative to Pseudeumeces cadurcensis (see Diagnosis above). Besides the features stated in the diagnosis, there is one additional difference – the dentary tooth number in Pseudeumeces cadurcensis is usually 17 (note that the tooth number in that taxon can range from 15 - 17; the holotype of the species [the left dentary AMNH FARB 241 A] is incomplete but the preserved portion bears seven teeth and at least five other empty tooth positions), whereas the dentary tooth number of Pseudeumeces kyrillomethodicus n. sp., based on material described here, is around 14 - 15. Although such small differences in tooth counts can be informative in some cases, it should be noted that the tooth number in lacertids (see e. g., Čerňanský & Syromyatnikova 2019), like virtually all lizards, should not be interpreted as absolute due to its variability. So, whether it seems that the tooth number of Pseudeumeces cadurcensis appears to be slightly higher than the new species, we cannot fully demonstrate it and we refrain from formally considering this feature as a diagnostic character. Besides the resemblance with Pseudeumeces cadurcensis discussed above, it should be noted also that the dentaries of Pseudeumeces kyrillomethodicus n. sp. slightly resemble those of Dracaenosaurus Pomel, 1846, in the following features (see Müller 2004; Augé 2005; Čerňanský et al. 2016 a, 2017): 1) dentary is a rather short, massive, and deep element; 2) the presence of a dorsally elevated posterior portion of the dentary; 3) the presence of the amblyodont dentition, where the posterior robust teeth are low and form blunt cylinders (this is more pronounced in Dracaenosaurus); and 4) the presence of striations on the tooth crown (note that the last two features are not unique to these two forms). However, there are some important differences between these two forms, where Pseudeumeces kyrillomethodicus n. sp. can be differentiated from Dracaenosaurus croizeti Gervais, 1848 - 1852, by the following combination of features (for Dracaenosaurus, see Müller 2004; Augé 2005; Čerňanský et al. 2016 a, 2017): 1) the dentary tooth number is around 14 - 15 rather than seven or eight; 2) the tooth size increases posteriorly, however, the largest tooth is the 4 th or 5 th one (counted from posterior), whereas the further posterior teeth decrease in size (to the contrary, the largest tooth in D. croizeti is usually the posteriormost one or sometimes the penultimate one); 3) the alveolar foramen, although its position can vary, is located further anteriorly (at the level between the 4 th and 5 th tooth positions in the holotype; counted from posterior) rather than at the level of the posteriormost tooth (or between last and penultimate tooth positions); 4) the facet for the anterolateral process of coronoid reaches around the level of the 4 th tooth position (counted from posterior) rather than terminating posterior to the tooth row; and 5) teeth (especially those in the posterior half of the tooth row) are anteroposteriorly compressed rather than mediolateraly compressed. Note that for Dracaenosaurus we follow recent workers and treat Dracaenosaurus sauvagei (Filhol, 1882) as a junior synonym of the type species Dracaenosaurus croizeti (see e. g., Augé 2005). In any case, the holotype dentary of D. sauvagei is different from that of Pseudeumeces kyrillomethodicus n. sp. and its tooth count is within the range of D. croizeti. In regards to our material, the specimen MNHN. F. QU 17169 (see Augé & Hervet 2009: fig. 1) deserves a comment. This specimen, which has been allocated to Pseudeumeces cadurcensis by Augé (2005) and Augé & Hervet (2009), has only 12 tooth positions instead of usual 16 - 17. Moreover, the dentary of this specimen appears to be robust rather than narrow. This would point to a huge level of variability. However, in MNHN. F. QU 17169, only the last posterior tooth is reduced and the coronoid reaches the level of this last posterior tooth position on both sides as it is typical, indeed, for Pseudeumeces cadurcensis (in contrast to our material described herein). In general, it can be expected stratigraphically that not identical but slightly similar forms of a lineage (the exact age of our material is unfortunately unknown, as is also that of the type material of Pseudeumeces cadurcensis) would exhibit a higher degree of morphological disparity reflecting the evolution through time than specimens collected from a single stratigraphic level. In those cases, it is of course difficult to add an exact border between such forms to distinguish taxa as units for science. However, we are convinced that all the above mentioned differences allow to erect a new taxon based on our type material. Even in extant herpetofaunas, lacertids include several morphologically cryptic species for which determination based on morphology can be even more difficult than the situation discussed here. Therefore, we consider the obvious differences in our material relative to the previously described forms as sufficient. Due to a high level of similarities of the currently known limited material with Pseudeumeces cadurcensis, we decided to allocate this new taxon Pseudeumeces kyrillomethodicus n. sp. to the same genus instead of erecting a new one. It should be further noted that the extinct genera Pseudeumeces, Dracaenosaurus, and Janosikia Čerňanský, Klembara, & Smith, 2016, have been recovered as sister taxa to the extant Gallotia Boulenger, 1916, from the Canary Islands, and all these taxa together with Psammodromus Fitzinger, 1826, form the clade Gallotiinae. This was firstly observed by Čerňanský et al. (2016 b, 2017), who also applied this revelation on the principles of the island rules, and later supported by Garcia-Porta et al. (2019) by their analyses based on a supermatrix relying on novel phylogenomic datasets. Therefore, we allocate Pseudeumeces kyrillomethodicus n. sp., as a member of Pseudeumeces, to the Gallotiinae as well. Recently described fossil material from the early Eocene (MP 8 - 9) French locality of Mutigny (Paris Basin) indicates that not only stem but also morphologically mod- ern-like (potentially crown or close to crown) lacertids were present on the European continent already in the early Eocene (Čerňanský et al. 2020). And later, lacertids were a rather diverse group during the Paleogene. The new taxon described herein, Pseudeumeces kyrillomethodicus n. sp., fully supports this high diversity and abundance of European Paleogene lacertids.
Georgalis, Georgios L., Čerňanský, Andrej, Klembara, Jozef (2021): Osteological atlas of new lizards from the Phosphorites du Quercy (France), based on historical, forgotten, fossil material. Geodiversitas 43 (9): 219-293, DOI: 10.5252/geodiversitas2021v43a9