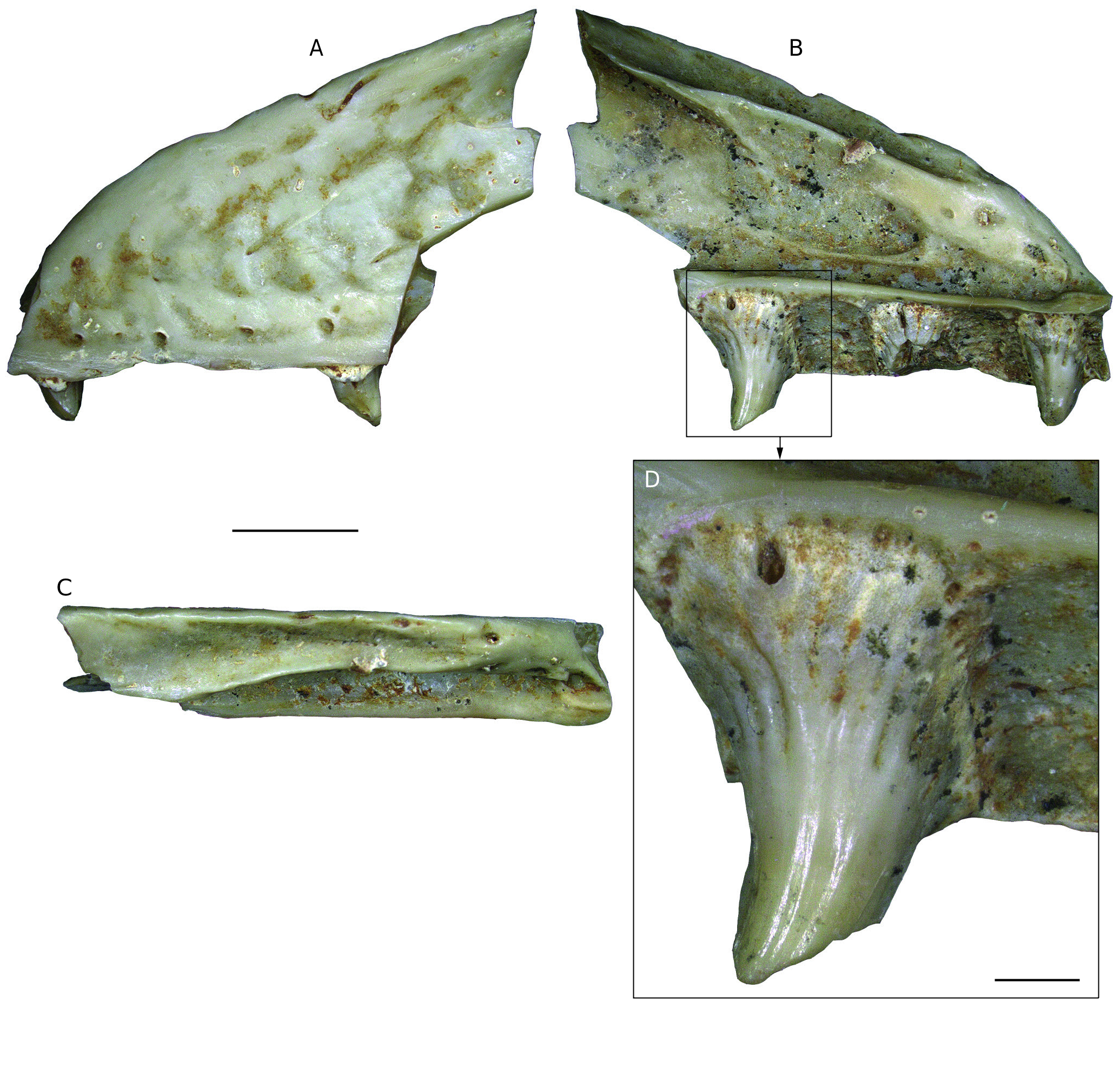
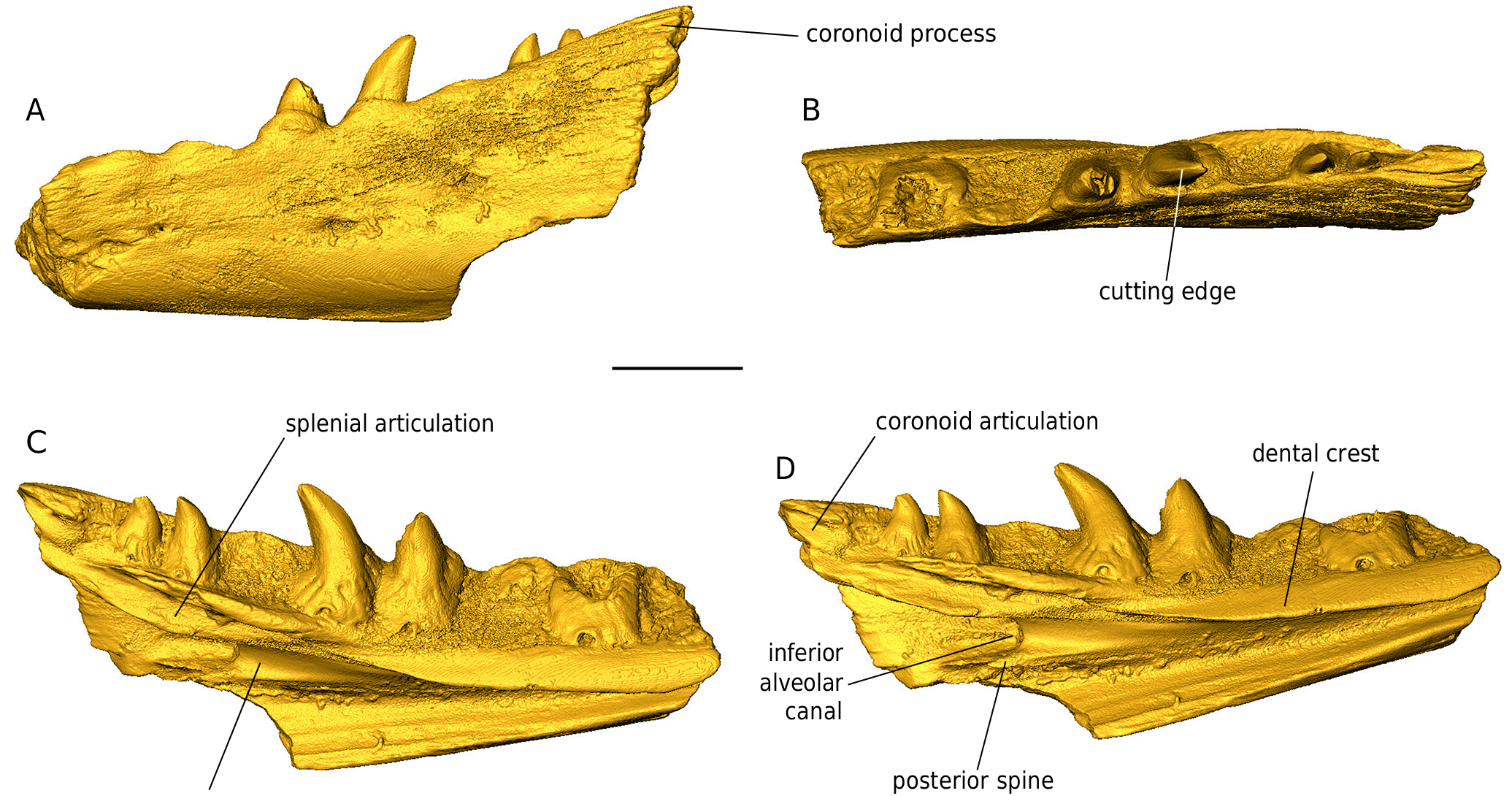
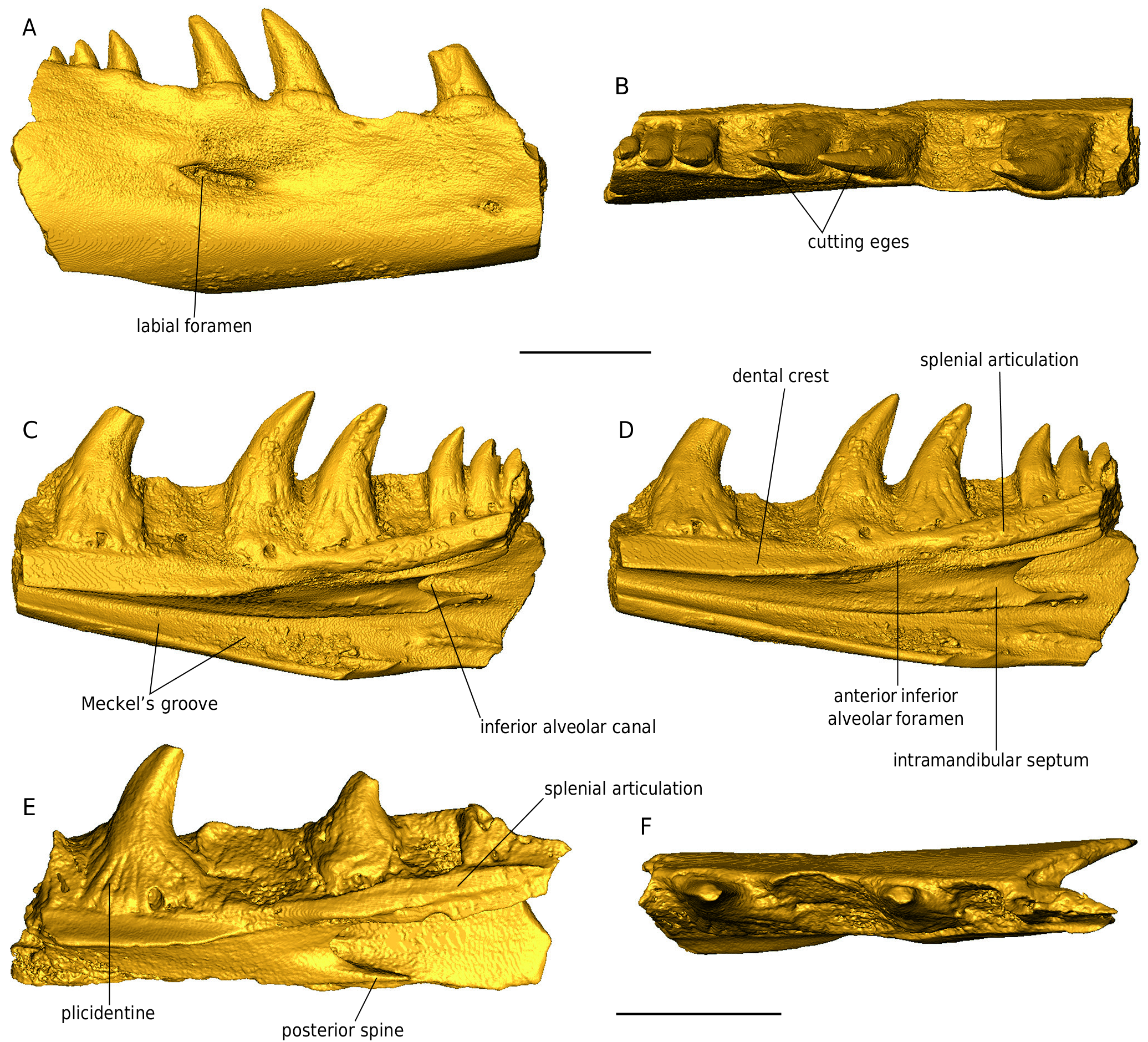
DESCRIPTION Maxilla NHMW 2019 / 0048 / 0002 (Figs 44; 45) The left maxilla NHMW 2019 / 0048 / 0002 is not fully preserved; its anterior half is well preserved, however, the anteriormost and the posterior portions are missing (Figs 44; 45). The external surface of the maxilla consists of large and low mounds divided by narrow grooves. Between the dorsal and ventral margins, the wall of the maxilla is distinctly concave. Immediately dorsal to the ventral maxilla margin, five labial foramina are preserved. The most distinguished feature of the dorsomedial surface of the nasal process (prefrontal process sensu Fejérváry 1935) of the maxilla is a shelf extending medially and running some distance ventral to the dorsal margin of the nasal process of the maxilla (Fig. 45 B, D, F). This shelf is the nasal crest which articulated with the nasal (a distinguished feature for Palaeovaranus). There is a deep sulcus between the medial margin of the nasal crest and the dorsal margin of the nasal process of the maxilla; the sulcus represents an articulating surface for the maxillary crest of the nasal. Immediately ventral to the mid-length of the nasal crest lies a semilunar impression in the medial surface of the nasal process of the maxilla; this is the articulating surface for the maxillary lamina of the prefrontal. Anteroventrally and anteriorly to the nasal crest, the medial surface of the maxilla is massively constructed; Fejérváry (1935) called this structure the basal torus (Fig. 45 B). Ventral to the semilunar impression and dorsal to the supradental shelf lies a large, elliptical depression in the medial wall of the nasal representing a lacrimal recess. Its anteriormost portion bears a foramen (Fig. 45 C). The supradental shelf is straight. Its dorsal surface bears a distinct sulcus for the nasolacrimal duct (Fig. 45 E). The anterior portion of the supradental shelf bears an elliptical and slightly dorsomedially inclined surface, an oblique inter- nasal lamella (Fig. 45 B). The maxilla immediately laterally to the supradental shelf is burrowed by the superior alveolar canal; its anterior and posterior openings are well preserved (Fig. 45 D, F). The bases of three teeth of the maxilla are well preserved. The bases are mesiodistally broad and bear well preserved basal striae, i. e., plicidentine (Fig. 44 B, D). The teeth are strongly curved distally and slightly lingually. The distal cutting edges are well developed. Dentaries (Figs 46 - 49) All three dentaries are incomplete, missing the anteriormost and posteriormost edges. The specimens NHMW 2019 / 0058 / 0054 and NHMW 2019 / 0058 / 0055 are relatively large and robust, with preserved maximum lengths 27.6 and 21 mm, while NHMW 2019 / 0058 / 0056 is smaller and slenderer, with a maximum preserved length of 18 mm. In all three dentaries, the maximum height is achieved at their posterior portions. NHMW 2019 / 0058 / 0054 bears four preserved teeth, one more tooth base, and three other empty tooth positions (Figs 46; 47), NHMW 2019 / 0058 / 0055 bears six preserved teeth and two other empty tooth positions (Figs 48 A, B; 49 A- D), and NHMW 2019 / 0058 / 0056 bears two preserved teeth, fragments of two other teeth, and two other empty tooth positions (Figs 48 C-E; 49 E, F). All teeth are subpleurodont, recurved, and rather pointed. Their bases are broad and are marked by the distinct presence of plicidentine. The latter feature is significantly most prominent in the anteriormost tooth of NHMW 2019 / 0058 / 0056 (Fig. 48 E). Teeth possess cutting edges, with this serration being most prominent in NHMW 2019 / 0058 / 0056. In the largest specimen (NHMW 2019 / 0058 / 0054), remains of the coronoid process are visible (Fig. 47 A). The coronoid articulation facet is preserved in NHMW 2019 / 0058 / 0054 (Fig. 47 D). Splenial articulation facets are visible in all specimens. The Meckel’s groove is open throughout the preserved lengths of all specimens. This structure becomes much widened posteriorly in all three dentaries, whereas in the anterior half of these specimens it is partially “ blanketed ” (but not closed) by the dental crest. In the posterior portion of the two largest dentaries, the characteristic alveolar canal (foramen dentale pro introitu partis alveolaris inferioris nervi mandibularis sensuFejérváry 1935) is prominent, delimiting a large longitudinal groove, which in NHMW 2019 / 0058 / 0055 extends and is visible also in its anterior edge. The posterior spine is thin, with its termination being acute. The intramandibular septum is wide (Fig. 47 C). In lateral view, several dorsoventrally elongated labial foramina pierce the dentaries’ surfaces, situated across irregular intervals, but otherwise these surfaces are relatively smooth (Figs 46 A; 47 A; 48 A, C; 49 A). Presacral vertebrae (Figs 50 - 53) The size of the presacral vertebrae varies, with centrum lengths ranging between 7.0 and 10.8 mm (Appendix 1). There are usually two distinct ridges in the ventral surface of the centrum that commence at around the level of the cotyle and terminate slightly prior to the condyle (Figs 50 - 53). In certain large specimens (e. g., NHMW 2019 / 0058 / 0005) a distinct deep groove is clearly delimited between these two ridges. Subcentral foramina are usually present and can be, at certain specimens, rather prominent (e. g., NHMW 2019 / 0058 / 0001; Figs 50 - 53). The neural spine is high in lateral view and is posteriorly inclined. The neural spine mostly develops and augments in height in the posterior half of the neural arch, but nevertheless, its base crosses, in all specimens, the entire midline of the neural arch, in the shape of longitudinal ridge. In certain large specimens, a distinct dorsal thickening of the neural spine is obvious (e. g., NHMW 2019 / 0058 / 0001; NHMW 2019 / 0058 / 0003; NHMW 2019 / 0067 / 0001). The neural arch is vaulted (or at least moderately vaulted) in all specimens. Foramina on the neural spine can be present (e. g., NHMW 2019 / 0058 / 0001). The cotyle is rather depressed; in fact, it appears to be exceedingly depressed in certain small-sized specimens (e. g., NHMW 2019 / 0058 / 0002; Fig. 52 A) but also in certain rather large ones (e. g., NHMW 2019 / 0058 / 0003; Fig. 53 A). In certain specimens, the cotyle is rather anteroventrally directed, so that its dorsal level can be observed even in ventral view of the vertebra (e. g., NHMW 2019 / 0058 / 0005; NHMW 2019 / 0058 / 0006), a condition reminiscent of varanids. In certain specimens (NHMW 2019 / 0058 / 0001; NHMW 2019 / 0058 / 0005), distinct anocotylar foramina are present above the cotyle. The synapophyses are massive and elongated. Sacral vertebrae (Figs 54 - 56) Two of these specimens (one isolated [NHMW 2019 / 0058 / 0029] and two articulated sacral vertebrae [NHMW 2019 / 0058 / 0028]) are rather large in size (Figs 54; 55; 56 A-F), especially the latter one, which exceeds in centrum length all the above described presacral vertebrae (see Appendix 1). The much smaller specimen NHMW 2019 / 0058 / 0047 is almost identical with the two other specimens, with their only major difference being this size contrast (Fig. 56 G-J); apparently it pertains to a much smaller (? younger) individual, however, any taxonomic implication for this size deviation cannot be excluded. Similarly to the presacral vertebrae, there are also in these specimens two distinct ridges in the ventral surface of the centrum, that commence at around the level of the cotyle and terminate slightly prior to the condyle. The centrum is convex in lateral view, with the ventral levels of both cotyle and condyle situated below the level of the ventral surface of the centrum. The neural arch is relatively vaulted. The neural spine develops mostly at the posterior half of the neural arch. The cotyle and condyle are depressed. “ Pseudozygosphenes ” and “ pseudozygantra ” are visible. The pleurapophyses extend much laterally and their distal termination is rather broad, so that in the two large articulated vertebrae (NHMW 2019 / 0058 / 0028), these elements from both vertebrae are united to each other (Fig. 55 D-F). A large fenestra is being formed by the unification of pleurapophyses of succeeding vertebrae. Also in this larger specimen, two distinct foramina are present below the neural arch in posterior view, inside the “ pseudozygantrum ” (Fig. 55 B). Caudal vertebrae (Figs 57; 58) These are extremely elongated and rather slender (Figs 57; 58), with the elongation becoming more prominent in the slenderer specimens (e. g., NHMW 2019 / 0058 / 0031; NHMW 2019 / 0058 / 0033), whereas the more robust ones tend to have a smaller centrum length but nevertheless wider vertebral centra (NHMW 2019 / 0058 / 0035; NHMW 2019 / 0058 / 0036; NHMW 2019 / 0058 / 0048). Their centrum lengths vary between 8.9 and 15.9 mm (see Appendix 1). The neural spine, when preserved, is rather high in lateral view, it augments in height rapidly but only at the posterior portion of the neural arch, and is inclined posteriorly (e. g., NHMW 2019 / 0058 / 0035; NHMW 2019 / 0058 / 0036). Nevertheless, the base of the neural spine extends much anteriorly in dorsal view, in the shape of a thin longitudinal ridge. The cotyle is large and elliptical. The prezygapophyses are dorsally inclined. The postzygapophyses are rather small. The pedicles for articulation with the chevrons are rather thick in the robust vertebrae and are located rather close to the condyle. A longitudinal groove runs throughout the centrum, originating posteriorly between the two pedicles and reaching anteriorly prior to the level of the cotyle. In the most robust specimens, this groove becomes rather thin in the anterior half of the centrum, while it is almost constantly wide in the slender, elongated specimens. There are no signs of autotomic septa. The transverse processes differ in size. In the most robust specimens, they occupy a larger surface at the lateral sides of the centrum, whereas in the more slender ones, they are rather thin. They extend laterally. In several specimens, there are “ pseudozygosphenes ” and “ pseudozygantra ” (sensu Hoffstetter 1969).
Georgalis, Georgios L., Čerňanský, Andrej, Klembara, Jozef (2021): Osteological atlas of new lizards from the Phosphorites du Quercy (France), based on historical, forgotten, fossil material. Geodiversitas 43 (9): 219-293, DOI: 10.5252/geodiversitas2021v43a9