Aequorea neocyanea
GBIF:190510757
ABOUT
Descriptions(7)
Export occurrence data
Darwin Core Archive (ZIP)
CLASSIFICATION
Taxonomic Classification Tree

MULTIMEDIA
Media Files(3)
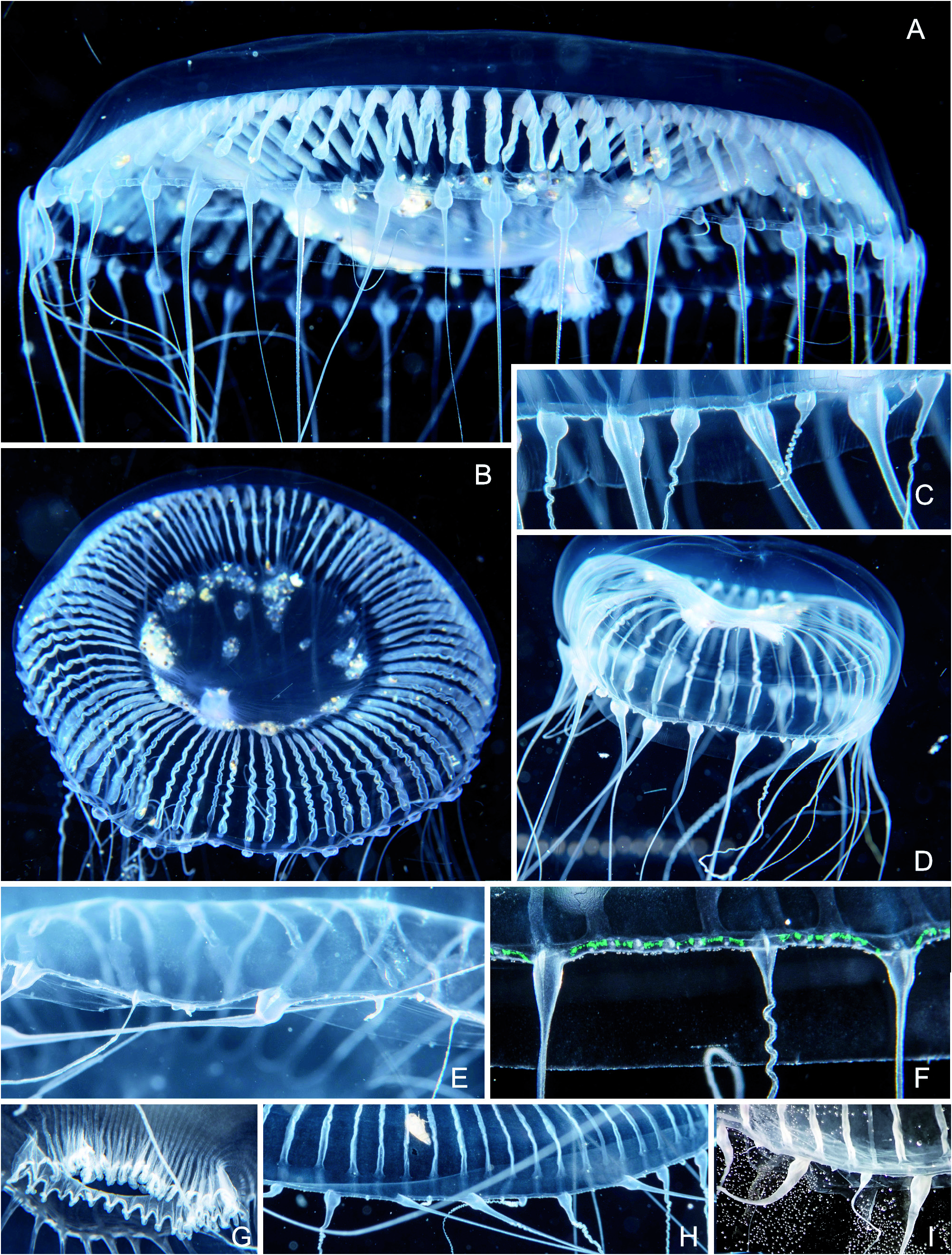
Fig. 38. Aequorea neocyanea new name. (A-C) BFLA4085, bell diameter 60 mm. (A) Oblique view from below. (B) Aboral view. (C) Bell margin with tentacles and their bulbs. (D) BFLA4083, bell diameter 42 mm, young animal with beginning gonad development. (E) BFLA4043, bell margin. (F) BFLA4236, bell margin, the green dots are not a pigment but likely due interference effects. (G) BFLA4236, mouth region with short fimbria. (H) BFLA3783, bell margin. (I) BFLA3783, tentacles after fixation in formalin.

Fig. 39. Aequorea forskalea. (A) Living medusa from Norway, bell margin seen from oral side. Photo courtesy of Luis Martell, sample HYPNO 772. (B) Preserved sample from the Mediterranean, MHNG-INVE-0055261.
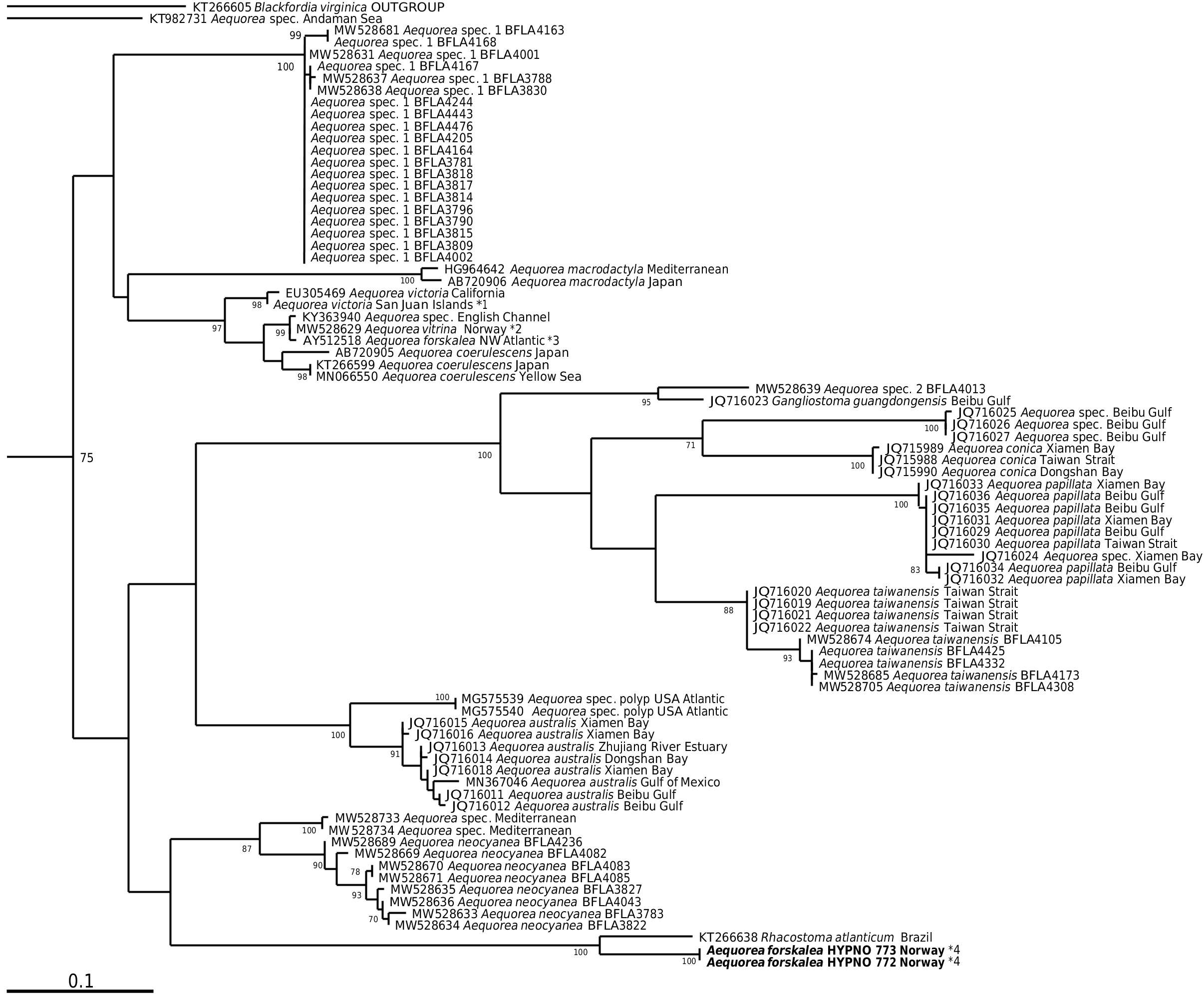
Fig. 37. 16S maximum likelihood phylogenetic tree of the genus Aequorea and related genera obtained with PhyML (GTR+G+I model) using about 600 bp positions of the mitochondrial 16S gene. Node-support values are bootstrap values of 100 pseudoreplicates (shown only if> 70%). Sequence labels start with the GenBank numbers (except for identical haplotypes) permitting the retrieval of more information. Red labels are new sequences from this study, for the taxa in bold either a voucher specimen or photos of it have been examined. Notes: *1) Unpublished, L. Leclère, pers. comm. *2) See Material & Methods. *3) Could be a misidentification. *4) Unpublished, A. Hosia & L. Martell, pers. comm., see Fig. 39A.
IMAGES