Pseudaegina rhodina
(Haeckel, 1879) Yes
GBIF:190510808
ABOUT
Descriptions(6)
Export occurrence data
Darwin Core Archive (ZIP)
CLASSIFICATION
Taxonomic Classification Tree

MULTIMEDIA
Media Files(3)
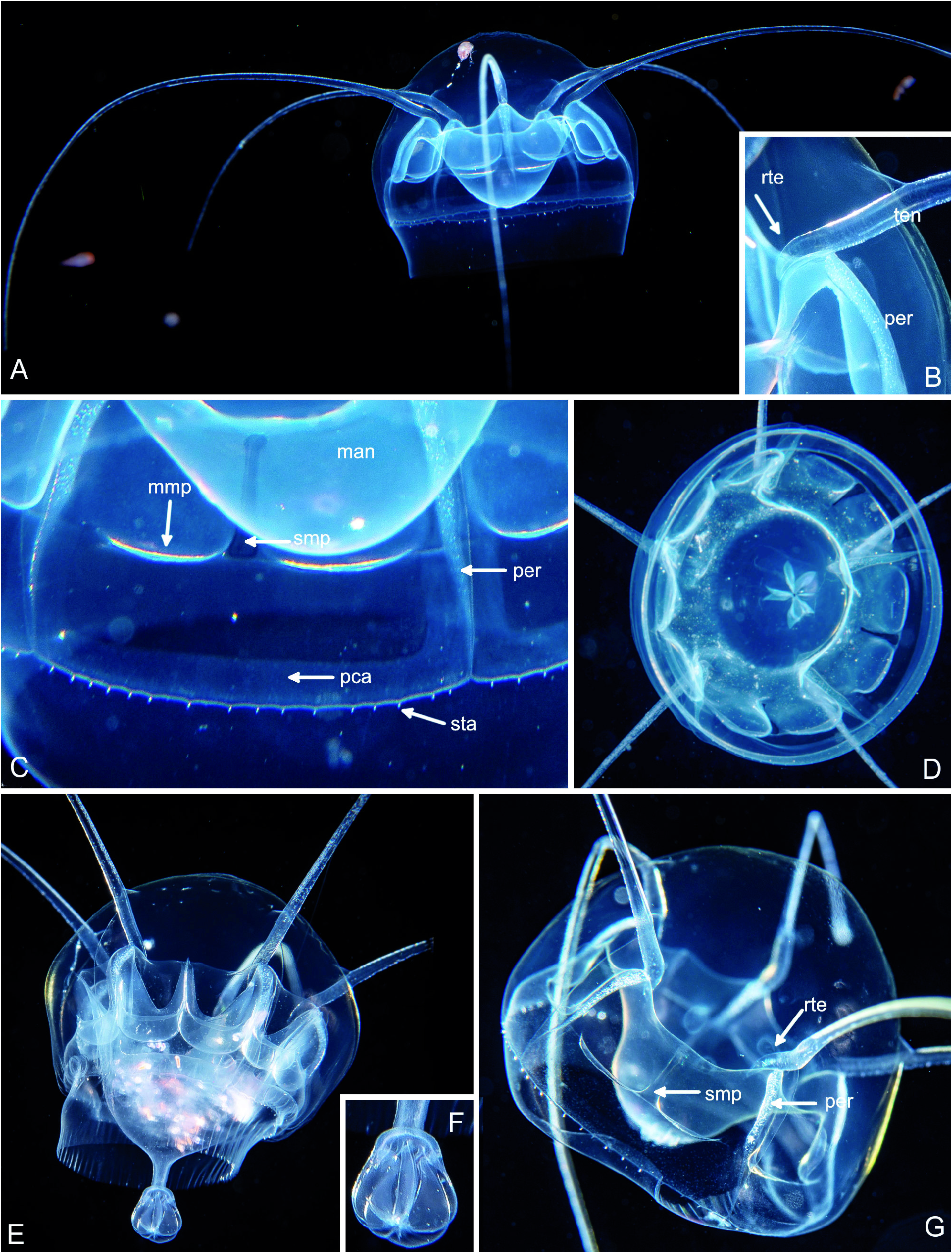
Fig. 51. Pseudaegina rhodina with 5 tentacles. (A-C) BFLA4015, size 18 mm. (D) BFLA4047, size 16 mm. (E-F) BFLA4067, size 20 mm. (G) BFLA4322, size 15 mm. (A) Lateral view. (B) Higher magnification of tentacle root (rte, arrow), note inflection towards below (= oral). (C) Details of bell margin. (D) Aboral view, note the (temporary) folding of mouth margin. (E-F) Medusa which has recently fed and its stomach is full of prey. Note the particular, most likely transitory, formation of a mouth tube and folded mouth margin, shown at higher magnification in F. (G). Lateral view. Abbreviations: man – manubrium/ stomach covering conical jelly cone, mmp – distal margin of manubrial pouch, pca – peripheral canal, per – peronium, rte – tentacle root, smp – septum in middle of manubrial pouch, sta – statocyst.

Fig. 52. Pseudaegina rhodina with 4 tentacles. (A) BFLA4134, size 9 mm, animal has semi-digested prey items in its gastric system including the peripheral canal system rendering it thus much more visible. (B) BFLA4120, bell size 9 mm, lateral view.

Fig. 48. Maximum likelihood phylogenetic tree of Narcomedusae obtained with PhyML (GTR+G+I model) and based on about 600 bp positions of the mitochondrial 16S gene. Node-support values are bootstrap values of 100 pseudoreplicates (shown only if> 70%). Sequence labels start with the GenBank numbers (except for identical haplotypes) permitting the retrieval of more information. Some proveniences were obtained from Lindsay et al. (2017) or through personal communications. Red ODEHOV DUH QHZ VHTXHQFHV IURP WKLV VWXG\. &RORXUHG ER[HV LQGLFDWH IDPLO\ DI¿OLDWLRQV DFFRUGLQJ WR WKH FXUUHQWO\ XVHG V\VWHP. (") 'HQRWHV SRVVLEOH PLVLGHQWL¿FDWLRQV.
IMAGES