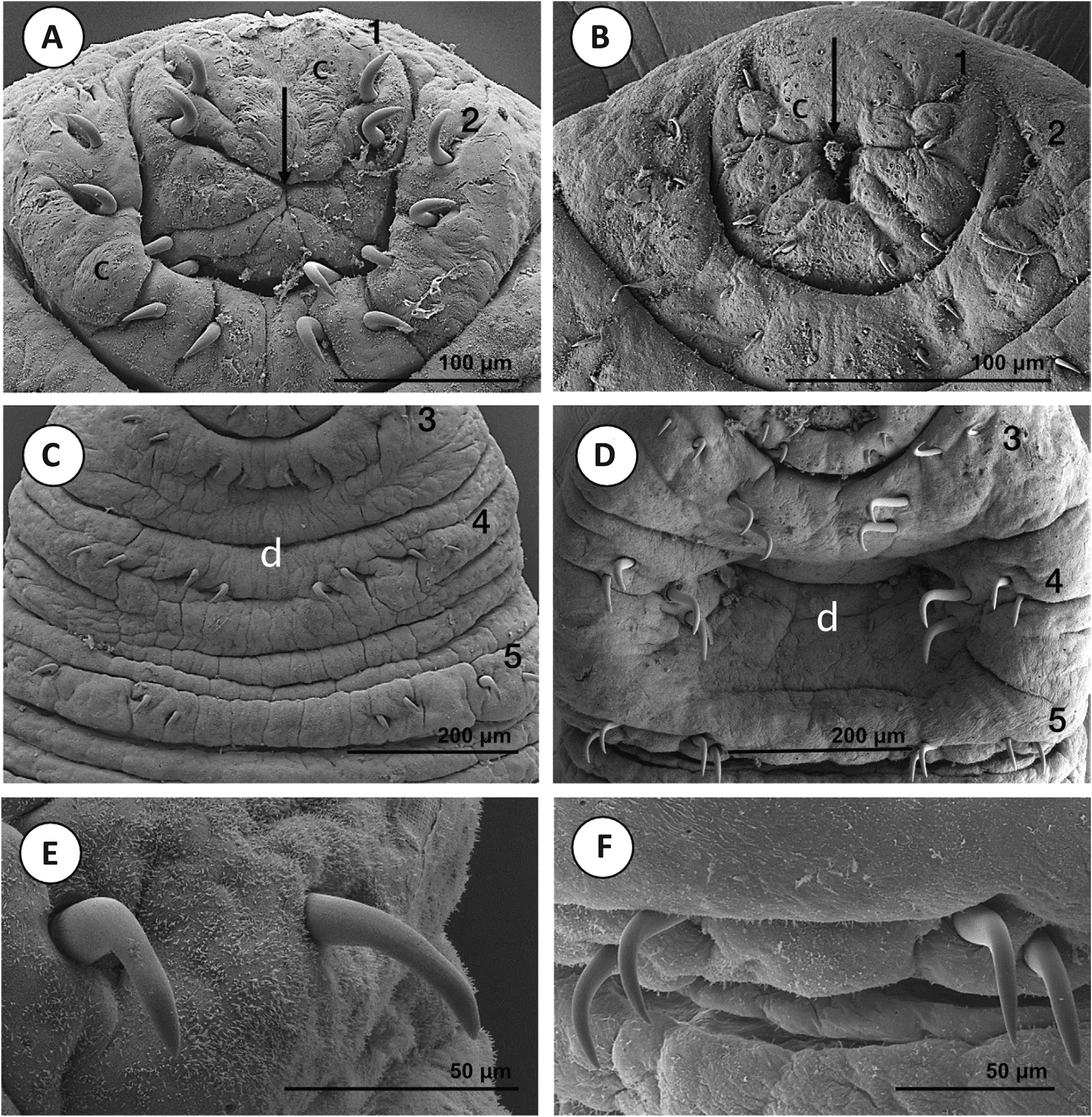
The present evidence suggests that A. peledina from Alaska is distinct, to some degree, from European samples. However, Siberia and the Russian Far East have not been adequately sampled genetically for A. peledina. Accordingly, it is difficult to determine the genetic variability and population structuring of this species. Coupled with increased taxon sampling, additional genetic sampling of Alaskan populations could help to indicate whether they are a unique species or population. Sampling of quickly evolving nuclear loci or, ideally, next generation sequence data (e. g. RADSeq) would be useful for determining whether gene flow exists between the Alaskan and Nordic localities. Unfortunately, COI, the most common marker for determining differences between leech species and populations (de Carle et al., 2017; Tessler et al., 2018 c; Mack et al., 2019), and additional nuclear loci did not amplify for these samples, potentially leading to some issues with missing data. Furthermore, given that no external morphological differences were noted between samples of Nordic and Alaskan A. peledina, we refrain from formal species or population delimitation analyses at this time. Nevertheless, the fact that the Alaskan population is sister to, and genetically divergent from, the Nordic samples suggests that this is not an invasive or nonnative species that was translocated only in recent times by humans, which would have been plausible given that the first records of this species in Alaska came from the 1970 s (Holmquist, 1974; Hauck et al., 1979) and that it has not officially been reported since then, despite the clear importance of these American animals. The Nordic populations are fairly similar genetically, despite being sampled from multiple (albeit geographically close) countries. The maximum genetic distance at the COI locus is 1.52 %, which is below the average value (~ 2.4 %) typically reported for species of Hirudinea (Kvist, 2015; de Carle et al., 2017; Anderson et al., 2020; Mack et al., 2019; Iwama et al., 2019). However, the countries sampled are all in relatively close proximity. It would be most useful to add samples from central and eastern Russia. Unfortunately, a 12 S sequence for A. peledina in the Baikal region of Russia from a recent publication was not made publicly available (Bolbat et al., 2019). MORPHOLOGY Our morphological examination and comparison of Paracanthobdella and populations of Acanthobdella help to characterize these species further. The scanning electron micrographs (Figs 6 – 9) and morphometry of facial hooks (N = 1280) help to accentuate the main external differences between the two species: (1) the presence or absence of a cup-shaped depression between rows of chaetae (anterior sucker); and (2) the chaetal dimensions and shape (Fig. 3; Table 3). The differences in both these characteristics become more notable as the species mature. The deep cupshaped anterior sucker, which is viewed as the most important distinguishing feature of P. livanowi, develops gradually through ontogeny from a flat state characteristic of juvenile individuals of the species. In A. peledina, the anterior end does not form a clearly separated sucker even in fully grown specimens, but a deep cavity appears between chaetae concomitantly with the growth of the animal. The shape of the chaetae differs between species: in A. peledina, chaetae are bent at a right angle and the breadth of the chaetae is similar in all rows, whereas in P. livanowi the angle is obtuse and chaetae in rows 4 and 5 have substantially greater breadth (Fig. 3; Table 3). The well-developed prostomium, which has been considered as another distinguishing feature of the species and the genus, was found to be less prominent and conspicuous than presented in previous studies (Epstein, 1987). Other studies have examined the internal morphology of these species (Bielecki et al., 2014); known differences from this work and others are summarized in Table 1. Acanthobdella peledina has the same morphology across the Nordic and Alaskan populations examined here and seems to be indistinguishable from Siberian populations (I. A. Kaygorodova & P. Świątek, unpublished scanning electron microscopy data) in other studies (Kaygorodova et al., 2012). Nevertheless, although we did not find differences between A. peledina from Alaska and Eurasia, it is entirely possible that detailed internal examinations might unearth differences, given that these populations appear to be divergent genetically. CONCLUSION AND FUTURE DIRECTIONS Our results help to shed light on the hook-faced fish worms (Acanthobdellida): an ancient lineage that is most closely related to leeches, and demonstrate that Acanthobdellida species and populations have diverged fairly recently. It is even possible that there are multiple species within A. peledina. Specifically, the American and Nordic populations appear to be distinct genetically and are likely to be isolated reproductively. However, there are important gaps to fill in the knowledge of the populations of this species before definitive action is taken on determining whether they represent the same species. Those gaps are as follows: (1) adding specimens from localities for central and eastern Russia; (2) obtaining additional genetic data (i. e. COI and additional nuclear data) for Alaskan samples; and (3) looking for internal morphological differences between populations.
Carle, Danielle Božena De, Gajda, Łukasz, Bielecki, Aleksander, Cios, Stanisław, Cichocka, Joanna M., Golden, Heidi E., Gryska, Andrew D., Sokolov, Sergey, Shedko, Marina Borisowna, Knudsen, Rune, Utevsky, Serge, Świątek, Piotr, Tessler, Michael (2022): Recent evolution of ancient Arctic leech relatives: systematics of Acanthobdellida. Zoological Journal of the Linnean Society 196: 149-168, DOI: 10.1093/zoolinnean/zlac006