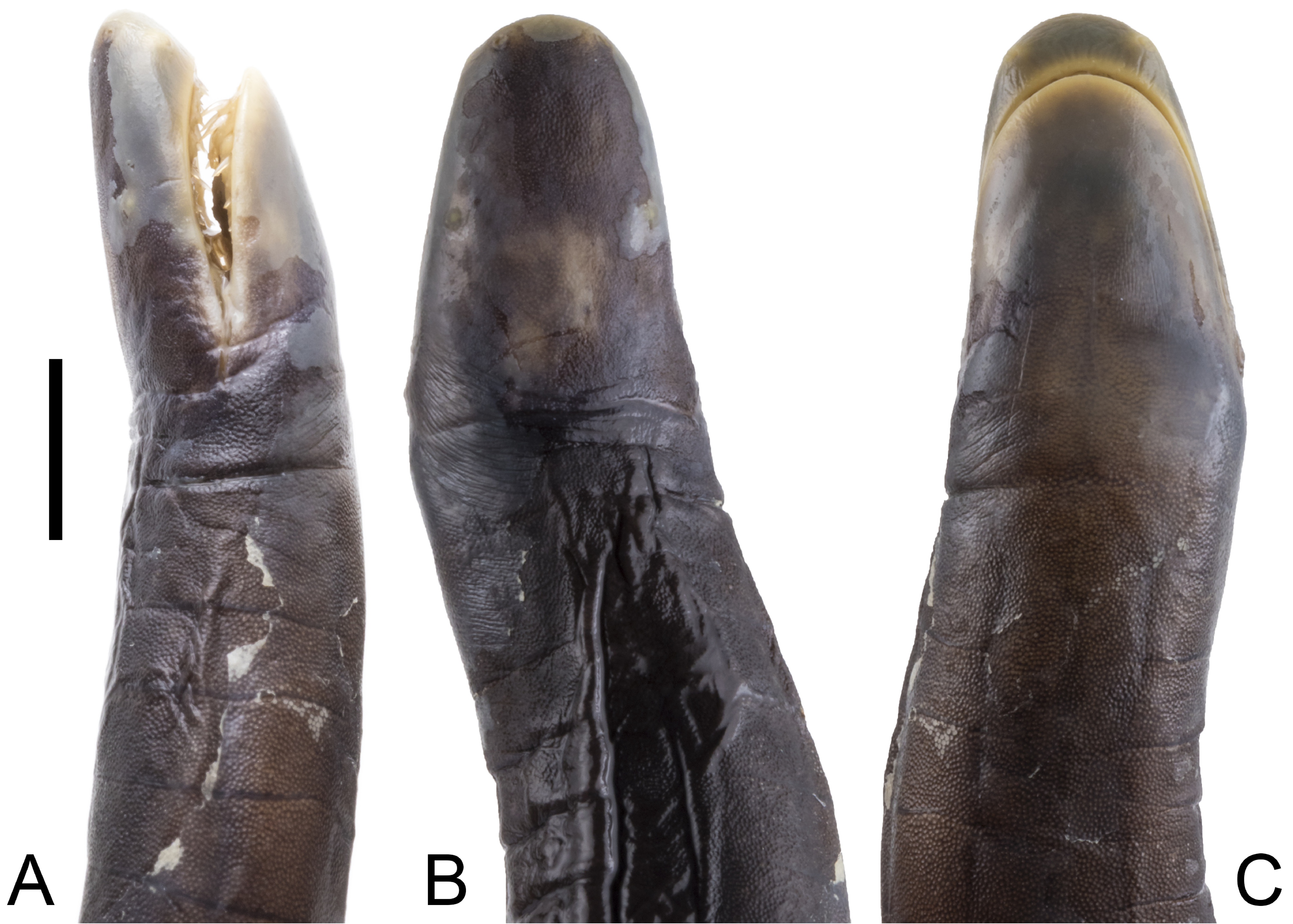
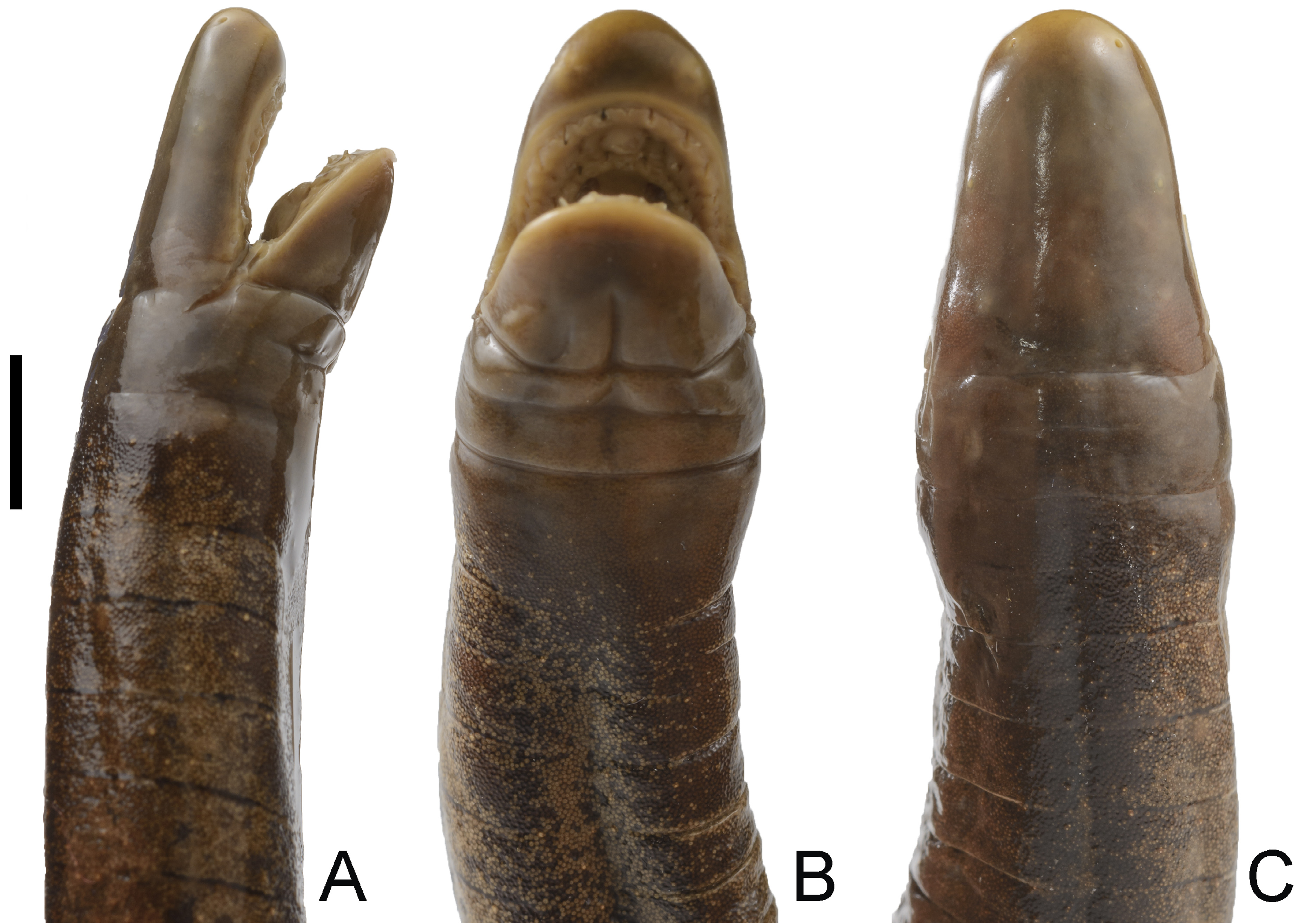
Diagnosis. Caecilia wilkinsoni sp. nov. differs from C. atelolepis, C. attenuata Taylor, 1968, C. caribea, C. corpulenta Taylor, 1968, C. crassisquama Taylor, 1968, C. degenerata, C. guntheri, C. inca Taylor, 1973, C. macrodonta, C. occidentalis, C. orientalis, C. pachynema (Günther, 1859), C. pulchraserrana, and C. subdermalis in having (vs lacking) secondary grooves. Caecilia wilkinsoni sp. nov. differs from congeners that have secondary annular grooves as follows. Caecilia disossea Taylor, 1968 (216 – 262 primary grooves and 16 – 34 secondary grooves) is the only member of the genus that has more primary and fewer secondary grooves than C. wilkinsoni sp. nov. (190 – 195 primary grooves and 51 – 71 secondary grooves). Caecilia aprix also has more primary grooves (234) but an overlapping secondary groove count (58) with the new species. Caecilia bokermanni (180 – 192 primary grooves and 15 – 21 secondary grooves), C. gracilis Shaw, 1802 (183 – 204 primary grooves and 11 – 21 secondary grooves), C. occidentalis (186 – 221 primary grooves and 0 – 15 secondary grooves), and C. thompsoni (187 – 240 primary grooves and 26 – 42 secondary grooves) have overlapping counts of primary grooves and fewer secondary grooves than the new species. Caecilia wilkinsoni sp. nov. has more primary annular grooves than C. abitaguae Dunn, 1942 (137 – 148 primary grooves and 0 – 5 secondary grooves), C. albiventris Daudin, 1803 (144 – 147 primary grooves and 45 – 53 secondary grooves), C. antioquiaensis (171 primary grooves and 4 secondary grooves), C. armata Dunn, 1942 (186 primary grooves and 92 secondary grooves), C. dunni Hershkovitz, 1938 (124 primary grooves and 67 secondary grooves), C. epicrionopsoides (139 – 163 primary grooves, 21 – 40 secondary grooves), C. flavopunctata Roze & Solano, 1963 (155 primary grooves and 27 secondary grooves), C. guntheri (111 – 132 primary grooves and 0 – 10 secondary grooves), C. isthmica (131 – 147 primary grooves and 12 – 21 secondary grooves), C. leucocephala (118 – 142 primary grooves and 17 – 45 secondary grooves), C. mertensi Taylor, 1973 (142 primary grooves and 48 secondary grooves), C. museugoeldi Maciel & Hoogmoed, 2018 (152 primary grooves and 26 secondary grooves), C. nigricans (157 – 189 primary grooves and 32 – 62 secondary grooves), C. perdita (139 – 152 primary grooves and 64 – 83 secondary grooves), C. subnigricans (151 – 161 primary grooves and 17 – 31 secondary grooves), C. subterminalis Taylor, 1968 (170 primary grooves and 16 secondary grooves), C. tentaculata (122 – 137 primary grooves and 30 – 42 secondary grooves), and C. volcani Taylor, 1969 (112 – 124 primary grooves and 14 – 32 secondary grooves). Caecilia nigricans (159 – 195 primary grooves, 27 – 65 secondary grooves, length in width 32.6 – 80.4 times) — another species from the Pacific lowlands and Cordillera Occidental of Colombia — has overlapping counts of primary and secondary grooves, and attenuation index values with C. wilkinsoni sp. nov. (190 – 195 primary grooves, 51 – 71 secondary grooves, length in width 65.6 – 74.7 times), but C. nigricans has a terminal shield or ‘ ’ cap’ ’ at the posterior end of the body that is barely interrupted dorsoventrally by the last few grooves of the body vs an entirely segmented terminal portion of the body in the new species, meaning that only the small disk and vent are free of grooves in C. wilkinsoni sp. nov. (Figs. 3, 6). Dermal scales obtained at the posterior end of the body of Caecilia nigricans are subcircular vs subrectangular in C. wilkinsoni sp. nov. Moreover, in life, Caecilia nigricans is mostly blue but C. wilkinsoni sp. nov. is mostly black. The new species is readily differentiated from C. tenuissima by having many more secondary grooves (51 – 71 vs 9). In addition, the new species has a head that is narrower than its mid-body width (vs noticeably wider than the body in C. tenuissima Taylor, 1973: 221, fig. 32); though in our experience a head much wider than mid-body is a rare condition within the genus, and the holotype of C. tenuissima appears to be somewhat desiccated from available photographs, suggesting this condition might be artefactual. Another difference between the species might be that in C. tenuissima the eye is not visible externally, while in C. wilkinsoni sp. nov. the eye is visible through translucent epidermis. There are no transverse grooves on the collars of C. tenuissima according to the original description (Taylor, 1973) but C. wilkinsoni sp. nov. has well-defined transverse grooves present on both collars. Squamation also differs between these two species, in that C. tenuissima has dermal scales only towards the terminus while C. wilkinsoni sp. nov. has dermal scales throughout its body. Caecilia wilkinsoni sp. nov. (190 – 195 primary grooves, 51 – 71 secondary grooves, length in width 65.6 – 74.7 times) is superficially similar to C. thompsoni (187 – 240 primary grooves, 26 – 42 secondary grooves, length in width 62 – 100 times), an endemic species of the Magdalena Valley of Colombia and the adjacent Cordilleras Central and Oriental, as well as the largest caecilian in the World, achieving a notable total length of 1767 mm (Arredondo-Salgar, 2007). However, the new species has more secondary annular grooves than C. thompsoni (51 – 71 vs 26 – 42) and has small, flattened (not protruding) narial plugs on the tongue (vs protruding narial plugs in C. thompsoni).
Fernández-Roldán, Juan David, Lynch, John D. (2023): A new species of Caecilia Linnaeus, 1758 (Amphibia: Gymnophiona: Caeciliidae) from the Pacific lowlands of Colombia, with comments on the status of C. tenuissima Taylor, 1973. Zootaxa 5270 (2): 194-206, DOI: 10.11646/zootaxa.5270.2.2, URL: http://dx.doi.org/10.11646/zootaxa.5270.2.2