Animalia
Animals(+39)
GBIF:1
0
Descendants
0
Children
0
Species
0
Genera
0
Families
PROFILE
Species Profile
Characteristics
ABOUT
Descriptions(25)
We include details on diversity for Central American representatives of 10 arachnid orders, excluding hyperdivrse mites & ticks. Individual taxa (families, genus, species) can be explored under taxonomy tab, using the dropdown hierarchical menu on the left, either directly or starting by the links below.
AMBLYPYGI: http://arachnids.myspecies.info/taxonomy/term/12
ARANEAE: http://arachnids.myspecies.info/taxonomy/term/18
OPILIONES: http://arachnids.myspecies.info/taxonomy/term/20
PALPIGRADI: http://arachnids.myspecies.info/taxonomy/term/15
PSEUDOSCORPIONIDA: http://arachnids.myspecies.info/taxonomy/term/13
RICINULEI: http://arachnids.myspecies.info/taxonomy/term/14
SCHIZOMIDA: http://arachnids.myspecies.info/taxonomy/term/17
SCORPIONES: http://arachnids.myspecies.info/taxonomy/term/16
SOLIFUGAE: http://arachnids.myspecies.info/taxonomy/term/19
THELYPHONIDA: http://arachnids.myspecies.info/taxonomy/term/11
Key to Chalcovietnamicus species groups and species
1. Embolus screw-like, without large embolic disc (ED) hidden between palpal bulb and cymbium; retromargin of chelicerae with one large fissidentate tooth of four cusps (Logunov 2020: figs 5–6)......................................... C. naga
- Embolus not screw-like, with large embolic disc (ED) hidden between palpal bulb and cymbium; retromargin of chelicerae with one bicuspid tooth (Figs 141–146)................................................... 2 ( Chalcovietnamicus s. s.)
2. Body with three conspicuous golden setal bands on carapace and dorsal abdomen (Figs 105–109; 118–125); a large apical flag-like embolic apophysis (EA) present on the dorsal side of embolus (Figs 116–117, 133–134)....... 3 (vietnamensis- group)
- Body covered with dense khaki setae (Figs 1–11, 29–30, 41–48, 70–76, 88–94); dorsal side of embolus lacking large apical flag-like embolic apophysis (EA; Figs 60–65).................................................. 4 ( daiqini -group)
3. Males with golden setal bands on lateral sides of dorsal carapace (Wang & Li 2022: fig. 4C); embolus without distal retro-ventral keel (dk; Fig. 117); in females, accessory glands (AG) visible in dorsal view of vulva (Fig. 115).............. C. lii
- Males have no golden setal bands of scale setae on lateral sides of dorsal carapace (Figs 120, 124), embolus with distal retro-ventral keel (dk; Fig. 133); in females, accessory glands (AG) invisible in dorsal view of vulva (Fig. 136)... C. vietnamensis
4. Median part of embolus bent near 90° in ventral view (Figs 97, 101); median septum of epigynum obviously raised (Fig. 99); distance between spermathecae longer than diameter of each spermatheca (Figs 100, 104)............ C. weihangi sp. nov.
- Median part of embolus not bent near 90° in ventral view (Figs 141–146); median septum of epigynum not raised (Figs 17, 35, 54, 82); distance between spermathecae shorter or equal to diameter of each spermatheca (Figs 17, 36, 55, 83)........... 5
5. Embolus S-shaped in ventral view (Fig. 80); copulatory ducts (CD) shorter than diameter of spermathecae, turning of copulatory ducts very close to spermathecae (Figs 83, 87)............................................... C. terbakar sp. nov.
- Embolus not S-shaped, almost straight or weakly bent in ventral view (Figs 141–146); copulatory ducts not as above...... 6
6. Embolus concave at dorsal side (Figs 66–68); atria small, diameter of atria shorter than 1/3 of diameter of spermathecae (Figs 55, 58)............................................................................... C. marusiki sp. nov.
- Embolus not concave at dorsal side; atria relatively large, diameter longer than 1/3 of diameter of spermathecae.......... 7
7. Apical extension of embolus (aE) obviously backswept (Figs 60–61); copulatory ducts (CD) not obviously bent in ventral view (Fig. 17)..................................................................................... C. daiqini
- Apical extension of embolus (aE) not backswept (Figs 63–65); median part of copulatory ducts (CD) obviously bent (near 90°; Fig. 36) in ventral view................................................................. C. logunovi sp. nov.
Export occurrence data
Darwin Core Archive (ZIP)
GEOGRAPHY
Distribution Map
REGIONS
Geographic Distribution(4)
DATA
Occurrence Datasets
Common names used for this species across different languages and regions. Available in 13 languages and 9 countries. 4 preferred.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide. This taxon has names in 13 languages. 4 names preferred.
HIERARCHY
Child Taxa(50)
RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(46)
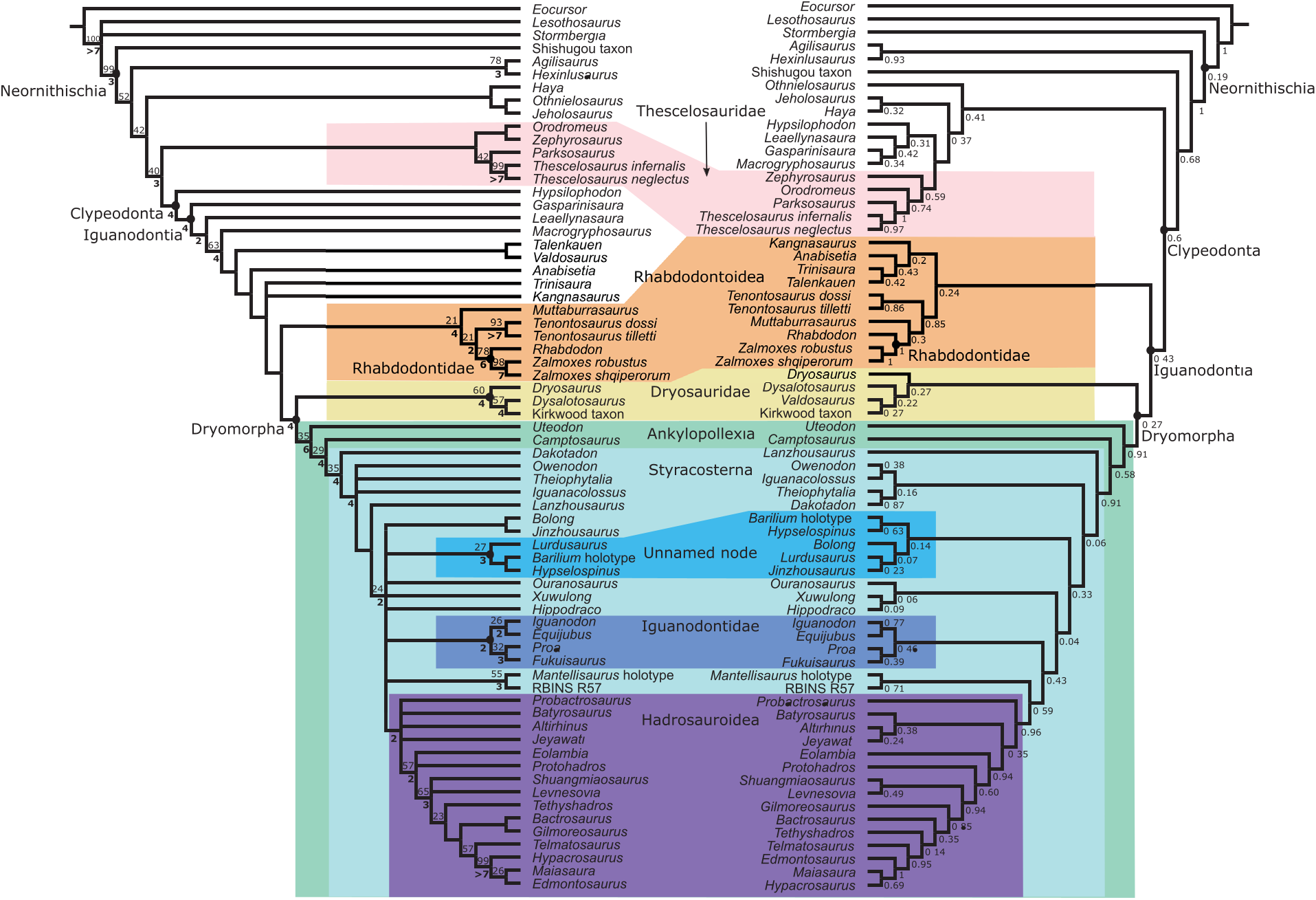
FIGURE 6. Parsimony (left) and Bayesian (right) trees plotted together. Strict consensus of 84 MPTs after pruning Oryctodromeus, Atlascopscosaurus, Planicoxa, Cumnoria, Cedrorestes, and NHMUK R28860. Jackknife values above 20 (with 10% chance of character removal) are shown above and to the left of their respective nodes. Bremer supports above one are shown below and to the left of their respective nodes, and bolded. CI=0.272, RI=0.634. Maximum clade credibility tree produced by Bayesian analysis showing posterior probabilities below and to the right of their respective node.
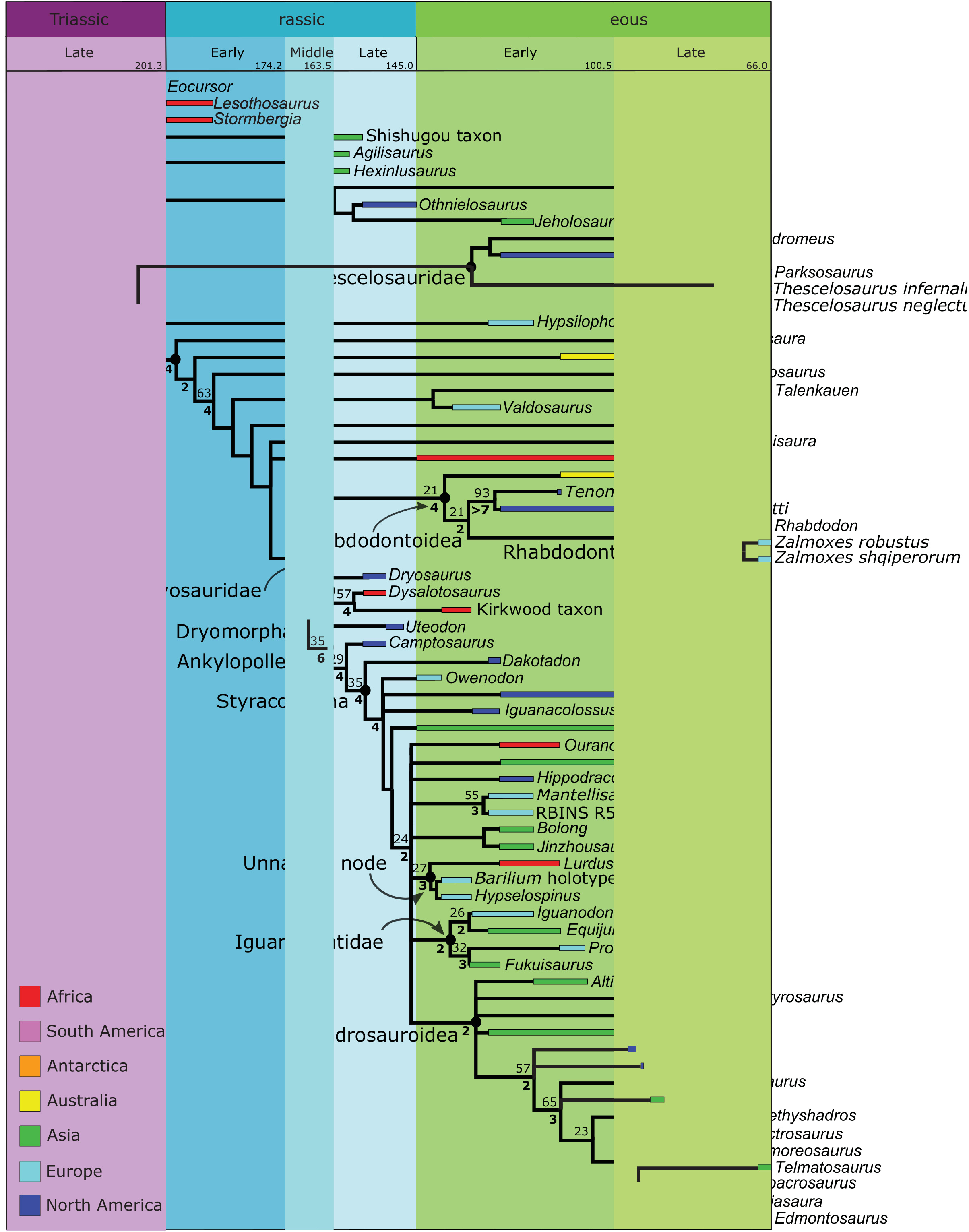
FIGURE 7. Time scaled parsimony tree, showing assigned age ranges and broad-scale geographic data. Jackknife values above 20 (with 10% chance of character removal) are shown above and to the left of their respective nodes. Bremer supports above one are shown below and to the left of their respective nodes, and bolded.
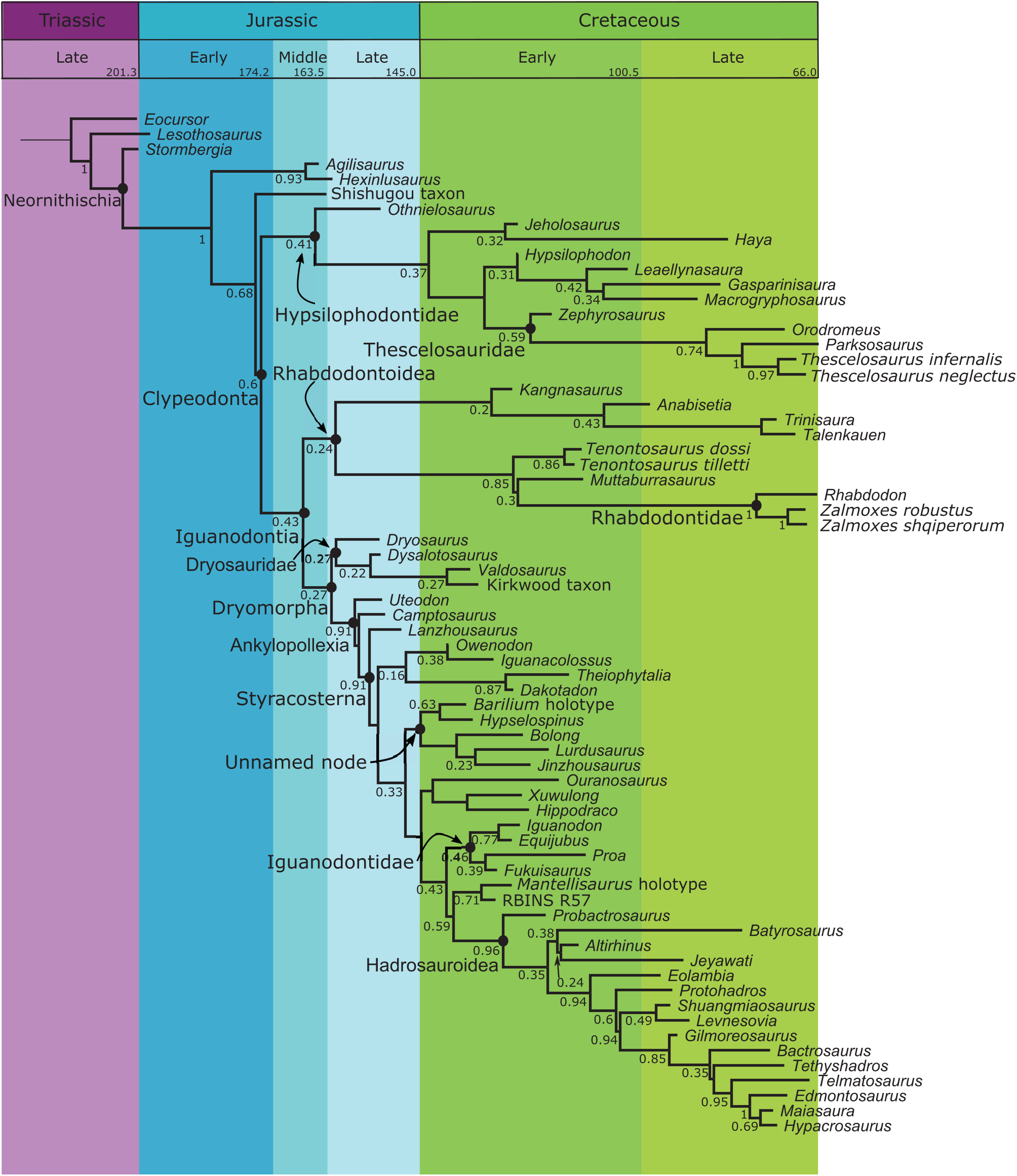
FIGURE 8. Maximum Clade Credibility tree produced by Bayesian analysis. Posterior probabilities are shown to the left of their nodes. The geologic timescale is shown across the top. Tips represent the average age found for each OTU across all sampled trees.
FIGURE 7. Time scaled parsimony tree, showing assigned age ranges and broad-scale geographic data. Jackknife values above 20 (with 10% chance of character removal) are shown above and to the left of their respective nodes. Bremer supports above one are shown below and to the left of their respective nodes, and bolded.
FIGURE 8. Maximum Clade Credibility tree produced by Bayesian analysis. Posterior probabilities are shown to the left of their nodes. The geologic timescale is shown across the top. Tips represent the average age found for each OTU across all sampled trees.
FIGURE 7. Time scaled parsimony tree, showing assigned age ranges and broad-scale geographic data. Jackknife values above 20 (with 10% chance of character removal) are shown above and to the left of their respective nodes. Bremer supports above one are shown below and to the left of their respective nodes, and bolded.
IMAGES
Gallery(46)
Occurrences with images
CITATIONS
References(6)
Margulis, L.; Schwartz, K.V. (1998). Five Kingdoms: an illustrated guide to the Phyla of life on earth. 3rd edition. Freeman: New York, NY (USA). ISBN 0-7167-3027-8. xx, 520 pp.
Margulis, L.; Schwartz, K.V. (1998). Five Kingdoms: an illustrated guide to the Phyla of life on earth. 3rd edition. Freeman: New York, NY (USA). ISBN 0-7167-3027-8. xx, 520 pp.
Parker, S.P. (ed). (1982). Synopsis and Classification of Living Organisms. McGraw-Hill, New York. 2 volumes.
Ruggiero, Gordon, Bailly, Kirk & Nicolson (2011) The Catalogue of Life Taxonomic Classification, Edition 2, Part A.
Ruggiero, M. A., D. P. Gordon, T. M. Orrell, N. Bailly, T. Bourgoin, R. C. Brusca, et al., 2015: Correction: A Higher Level Classification of All Living Organisms. PLoS ONE vol. 10, no. 6, e0130114.