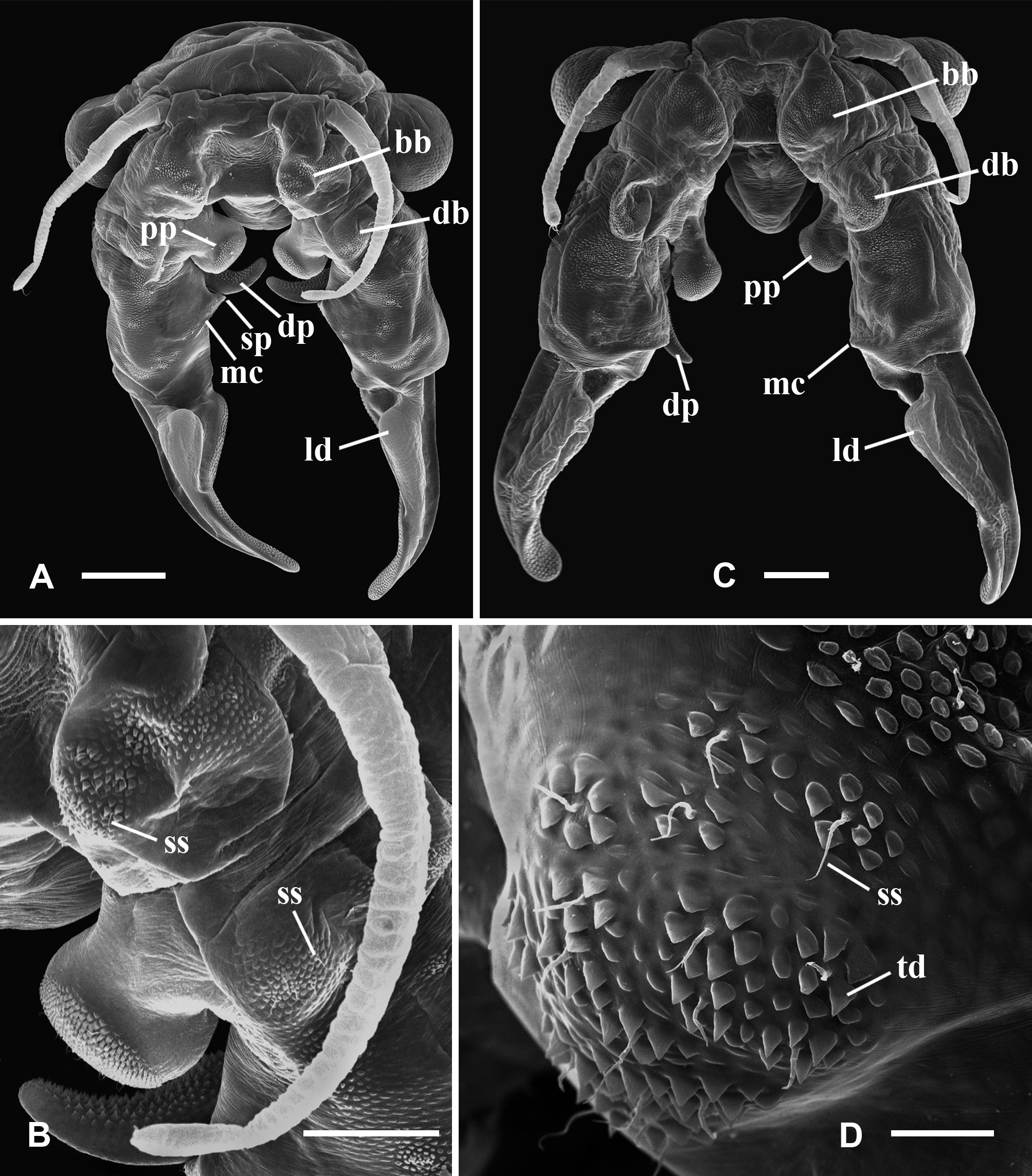
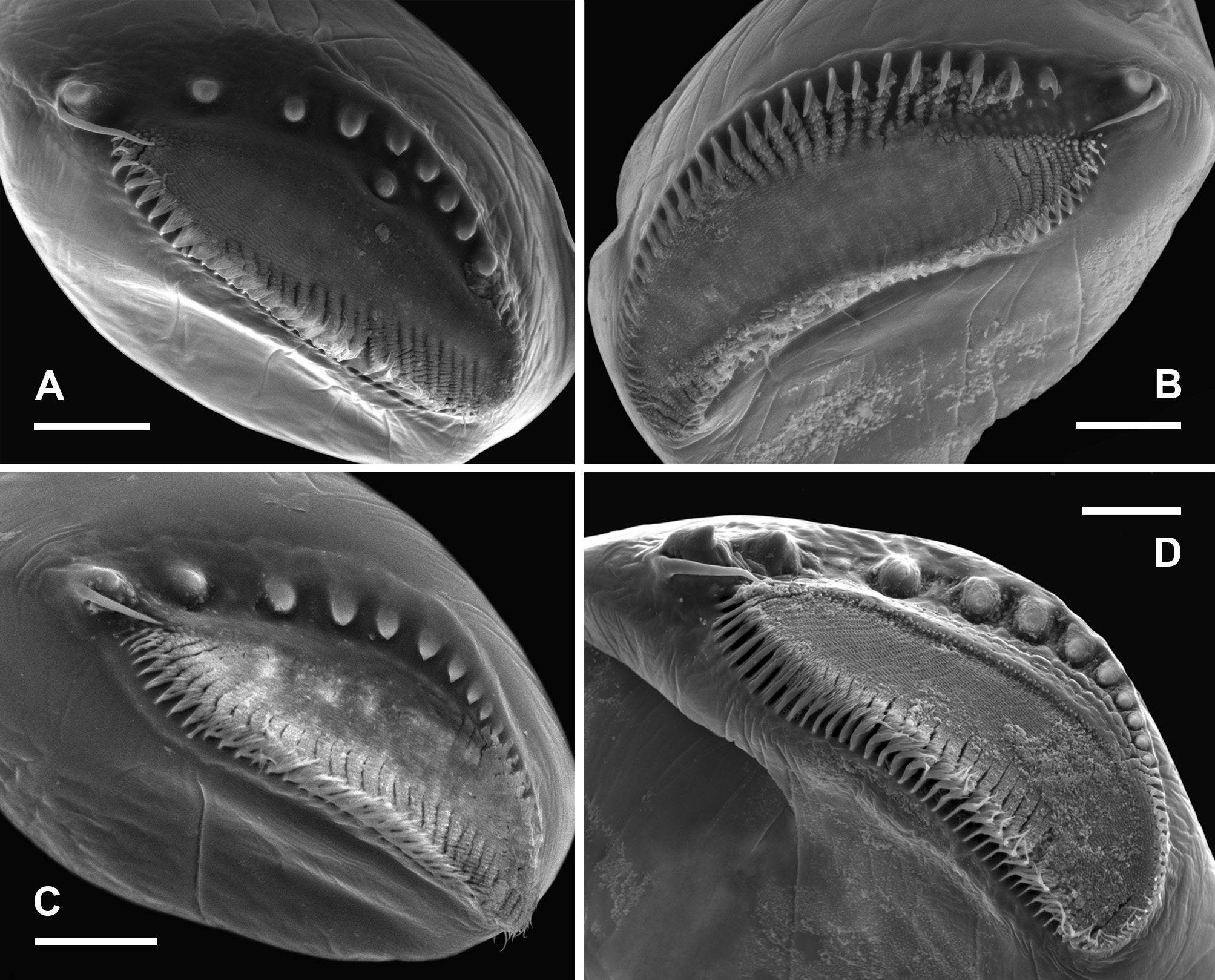
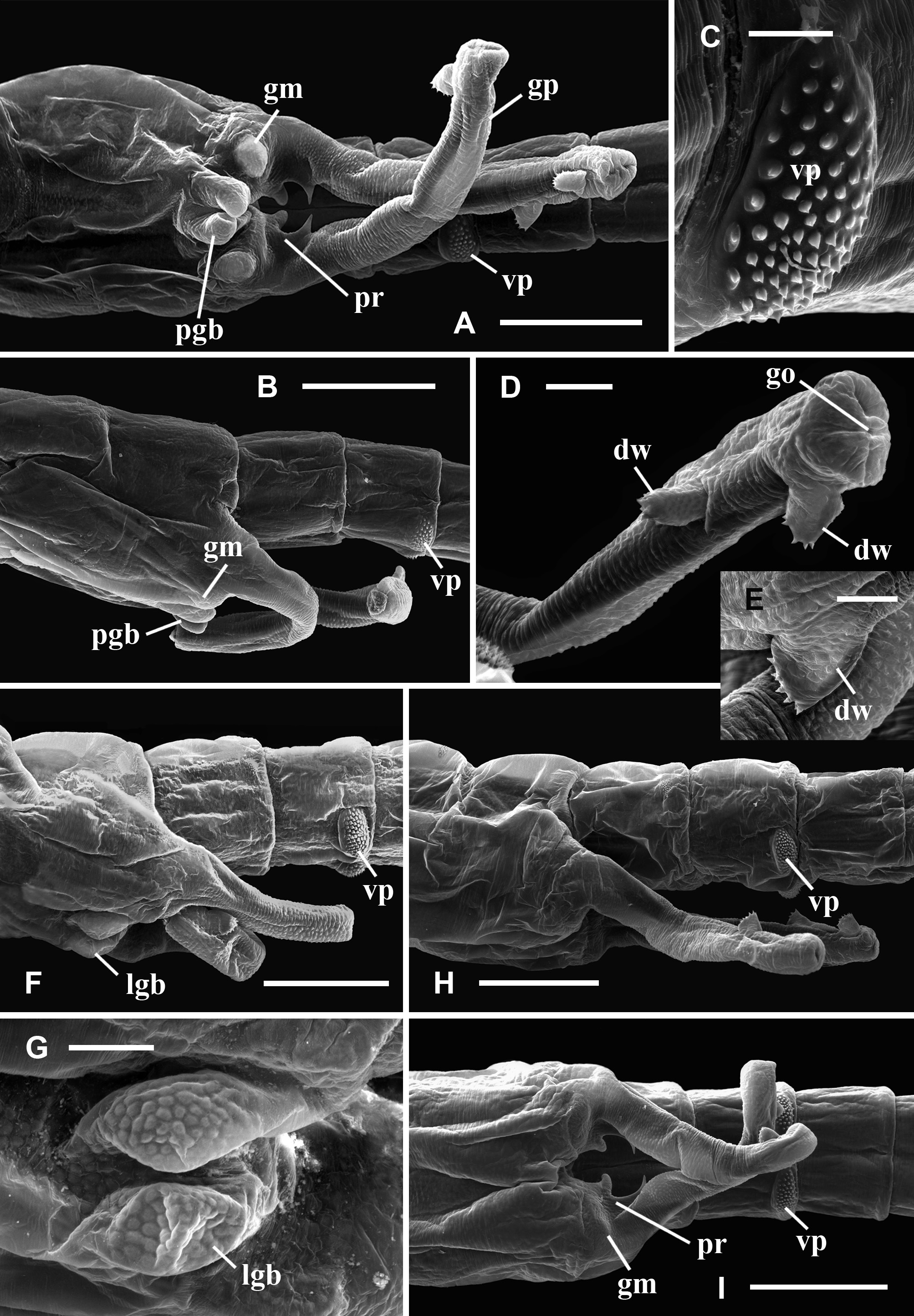
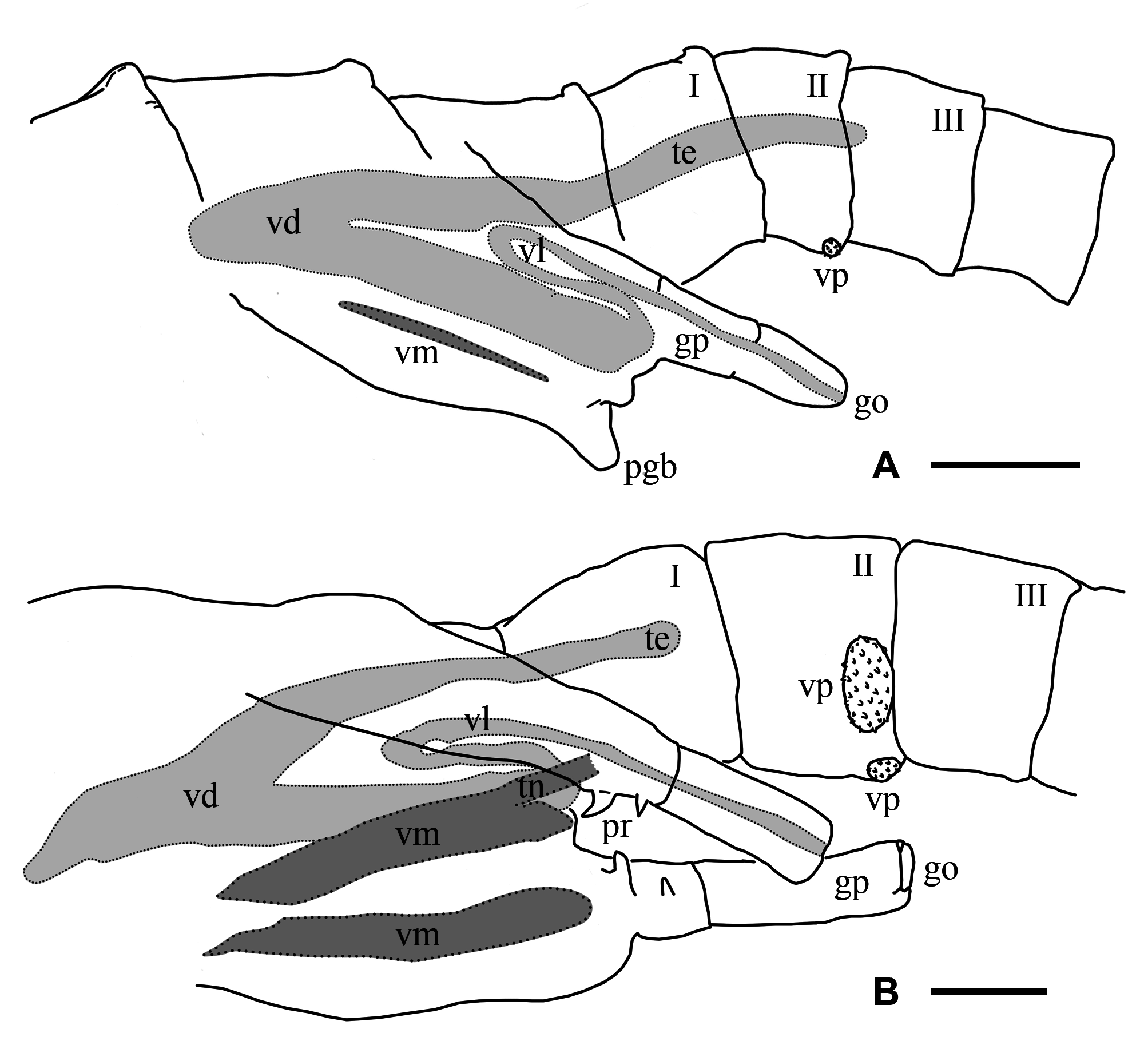
Description. Male. Cuticle ornamented with granulations throughout the whole body, mainly evident in the head, genital and abdominal segments, and gonopods. Compound eyes mean diameter: 0.6 mm (0.4 – 0.8 mm; n = 26), on average 1.5 times larger (1.4 – 2.0; n = 56) than in females (Tables 3, 4). First antennae partially multi-articulated, with incomplete rings (Fig. 2 C). Second antennae basal antennomere, with a basal swollen bulge (bb), on its anterior surface, near the medial edge (Figs. 2 C). Under SEM magnification, the bulge looks paved by small slightly convex cuticular plates, some of which rise and point anteromedially, forming triangular denticles arranged in circular groups encircling a central hyaline sensory seta. The basal antennomere bears on its anterior surface a more distal swollen bulge (db), similarly paved with convex small plates which do not rise as denticles, and scattered warty sensory areas surrounding a hyaline seta (Fig. 2 C); posteromedially at the same level, a spherical knob hangs from a peduncle as a slender pedunculate process (pp) distally dilated (Figs. 2 C; 5 A, D); the peduncle has a granulous cuticle, whereas the tip of the process is ornated with mesetiform elements tightly arranged, and interspersed setae (Fig. 5 A, D, E); more distally in the basal antennomere, and also protruding from its posteromedial surface, there is a digitiform process (dp) that may not be visible in anterior view of the head, remaining hidden behind the posterior face of the basal antennomere (Figs. 5 A, C, D), or its tip barely appears medially (Fig. 2 C); in any case, the digitiform process of each antenna does not reach the small process of its own side, nor projects medially until coming into contact with the digitiform process of the other second antenna. The digitiform process is ornamented with sharp denticles that are more developed on the surface facing the ornate tip of the pedunculate process; from this surface to both sides, denticles gradually flatten until disappearing, so that the cuticle appears almost smooth (Fig. 5 B). At the level of the origin of the digitiform process but in the medial surface of the antennomere there is a transverse oblique crest or medial crest (mc), ending posteromedially into a small process (sp) represented by a low mound (Fig. 5 A, C, D); the small process is ornamented with columnar grooved spines distally divided into several irregular points, scattered simple smooth spines and interspersed hyaline setae (Fig. 5 F). The distal antennomere bends from its anterior surface in posteromedial direction, narrowing distally as it curves, and culminating in a rounded spatulate tip (Fig. 2 C). A little beyond its proximal articulation, the anterolateral surface of the antennomere protrudes like a vertical globose triangular ledge (ld) with proximal base (Fig. 2 C), which narrows in distal direction and ends up at the distal third of the lateral edge of the antennomere. A wide rasp-like area covered with conspicuous mesetiform ornamentations, like those present in several Branchinecta species, is observed from the tip of the appendage, advancing on the posterior and medial surfaces of the distal antennomere. Mandibles asymmetric, showing no sexual dimorphism. In the left mandible (Fig. 6 C, D), starting at the posterior pole, a deepened posterodorsal region along the dorsal margin bears a row of teeth decreasing in size in anterior direction; the region extends up approximately to 3 / 4 of the margin, so that the teeth do not reach the anterior round- ed mandibular pole, and are gradually replaced by spiniform setae. The right mandible shows a similarly deepened posterodorsal region starting at the posterior pole but lacking teeth; instead, parallel rows of multibranched projections transversal to the major mandible axis, gradually develop; these rows progressively reach the level of the main masticatory surface, approximately at 1 / 4 of the dorsal margin. Both mandibles show at the posterior pole a tooth slightly stronger than those at dorsal margin of left mandible, associated with one long and thick seta (rarely two), curved inwards (Fig. 6 C, D). A reduced mandibular palp protrudes from a polygonal area in the convex side of the mandibular surface (Cohen 1991). Maxilla 1 with a strong ventral spine and 20 long setae; the spine is 1.5 – 2 times the width at the basal portion of the long setae, and barely surpasses in length this basal portion. Maxilla 2 with 8 – 10 plumose distal setae plus 2 – 3 anteriorly directed densely plumose setae, with setules all around. Thoracopods show no sexual dimorphism. Endopodite of thoracopods 1 to 2 – 3, transversely subrectangular with triangular apex; from thoracopod 3 or 4 to 7 the endopodite is triangular; in thoracopods 8 and 9 it is triangularoval; in thoracopods 10 and 11 it is oval. Lower edge: throughout the thoracopods 1 to 8, the 80 % of the lower edge of endopodite shows plumose setae; in distal direction the setae become long, slender, curved and pectinate, elongating to the apex of the lobe; in endopodites 9 and 10, these later setae become shorter and weaker; in endopodite 11 the plumose setae are followed in the distal direction by pectinate or bipectinate straight setae shortening to the apex. Medial edge: from the apex, the setae gradually decrease in length all along the medial edge of the endopodite. Here, the setae bordering the endopodite 1 are bipectinate and straight; those in endopodites 2 – 8 are pectinate and curved, though the most proximal ones are short and straight; in endopodite 9 and 10, the setae are pectinate but weaker, shorter and curved or straight; in endopodite 11 the setae are pectinate or bipectinate, short and straight; the proximal half of the medial edge is devoid of setae. Endites 1 and 2 throughout all the thoracopods showed the typical number and arrangement of anterior and posterior setae proposed by Linder (1941) for anostracans. In the 1 st pair of thoracopods the number of setae on the remaining endites was, in endite 3: 8 anterior setae plus a long spine and 3 posterior plumose setae; in endite 4: 7 – 8 anterior setae plus a long spine and 2 posterior plumose setae; in endite 5: 8 – 9 anterior setae and 2 posterior plumose setae. In pairs 2 to 11 the number of setae in the endite 3 was: 1 – 2 anterior seta plus a long spine and 3 posterior plumose setae (2 posterior setae, in pair 11); in the endite 4: 1 – 2 anterior seta plus a long spine and 2 posterior plumose setae; in the endite 5: 2 – 4 anterior setae and 2 – 3 posterior plumose setae. In addition, the thoracopods 1 – 10 show an oval epipodite with smooth margin, and a unique preepipodite with serrated margin, with or without a small marginal notch. In the pair 11, the epipodite is big, oval and margined with around 6 – 11 plumose setae, and the preepipodite is a triangular much-reduced lobe, with smooth margin and apex rounded. Genital and abdominal segments. Abdominal segment II (second postgenital segment) bearing, close to its posterior articulation, a pair of ovoid ventrolateral denticulate protuberances (vp), showing diverse degrees of separation from each other in different specimens (Fig. 7 F, H, I); these protuberances densely covered by denticles surrounding a seta (Fig. 7 C) seem to correspond to a hypertrophied pair of segmentary sensory areas. Similarly ornate but less prominent areas were observed in equal positions, near the posterior articulation of abdominal segments IV and VI. Besides, both genital segments present a conspicuous pair of dorsal warty sensory areas, also observed in several abdominal segments. Gonopods (gp) widely separated, extending ventrolaterally, with non-eversible portions partially visible in dorsal view; very long when completely everted, extending up to the abdominal segments III or the beginning of IV (Fig. 7 D, H, I). The rigid basal portion of gonopods bears a medial sclerotized projection (pr) with two processes (Fig. 7 I) as characteristic in the genus; in addition, more laterally in the basal portion there is a ventral pair of low granulous mounds (gm) (Fig. 7 I). In ventral view, near the distal edge of the second genital segment and between the rigid basal portions of both gonopods, there is a pair of low genital bulges (lgb), next to each other at the midline and distally rounded (Figs. 7 F, G); in lateral view they barely protrude or are almost indistinguishable (Fig. 7 H, I). Eversible portion of the gonopod very long, reaching the abdominal segment III or the beginning of IV (Fig. 7 H). Anterior or lateral longitudinal rows of denticles were not observed on this portion of the gonopod. Like in the other Branchinectidae, each eversible portion of the gonopods shows two dentate warts (dw) in its distal part (Fig. 7 D, E). Both warts are forwardly and obliquely directed, and bear 6 to 8 short marginal spines; one of the warts is medial and close to the tip of the gonopod and the other is dorsal and located at ¾ of the length of the everted portion of the gonopod (Fig. 7 D, H). The internal structure of the male genital segments in lateral or ventral view (Fig. 8 B), shows that each testis (te) is connected with the dorsal branch of a wide V-shaped portion of the vas deferens (vd) extending forwardly along both genital segments and turning to continue into a ventral branch that extends posteriorly throughout the genital region; each ventral branch narrows abruptly to become the thin loop portion of the vas deferens (vl) that enters in the partially everted or non-everted gonopod (gp), and then stretches when the penis is wholly everted. In ventral view, both branches of the vas deferens look superimposed, though they are hidden under a ventral muscle (vm) along the genital region, which is connected by means of a tendon (tn) with the non eversible portion of the gonopods. Testes reach abdominal segments I to III (Fig. 8 B). Uropods (or cercopods) length: 1.6 mm (1.1 – 2.6 mm; n = 26), longer than in females (male uropod / female uropod: 1.3 (0.9 – 1.5); n = 56), on average 3.2 times as long as the anal segment (or telson) (2.5 – 3.9; n = 26) (Table 3), divergent, uniformly provided with marginal plumose setae. Female. Cuticle ornamented with granulations, mainly evident in the head, genital and abdominal segments, and brood pouch. Compound eyes smaller than in male, mean diameter: 0.4 mm (0.3 – 0.5 mm, n = 30) (Table 3). First antennae as in male (Fig. 9 A). Second antennae presenting a strong conical medial projection (mp), in the inner surface, medially directed, giving the appendage a cheliform aspect (Fig. 9 C, E). The anterior surface of the antenna as well as the medial projection, including its tip, present warty sensory areas (sa) associated with a central seta (Fig. 9 E). Instead, the apical projection (ap) of the appendage lacks sensory areas (Fig. 9 E). Just proximal to the medial projection there is a medial bulge (mb) (Fig. 9 C, D), slightly rough, provided with hyaline setae, either like low (Bo- 35) or prominent (Bo- 18) warty areas. Mandibles as in male; no differences between sexes in the molar ornamentation are apparent. Maxilla 1, with a strong ventral spine and 24 – 25 long setae; the spine is almost four times the width of the basal portion of long setae, and barely surpasses in length that basal portion. Maxilla 2, with 4 – 5 plumose distal setae plus 2 anteriorly directed densely plumose setae, with setulae all around. Thoracopods showing the same morphology of lobes as in male. Particularly, the shape of endopodites is similar throughout the series, changing from subrectangular to triangular to oval (in pairs 10 and 11), and also both sexes agree in their setose armature bordering lower and medial edges. The number of anterior and posterior setae of endites 1 to 5 in thoracopods 1 – 11 is like in the male, differing in ± 2 setae. In thoracopods 1 – 10, the preepipodites present a serrated margin without a defined marginal notch; in pair 11, the preepipodite is a triangular lobe not as reduced as in the male, and the epipodite is big, oval and margined with around 9 – 10 plumose setae. Thoracic ornamentation. Dorsally poorly developed (Fig. 10 F, H), though it may vary even within the same sample (Fig. 10 G). Up to two pairs of evaginations, one dorsal and one dorsolateral, can be observed in each thoracomer 9 or 10 to 11. When the evaginations are present in segment 9, they are low, rounded, and one of the pairs may be missing; in segments 10 and 11 the dorsal ornamentations are rounded to conical, confluent in the midline, and the dorsolateral ones are usually low, globose or conical or may be missing. Thoracomer 12 (first genital segment) with a conspicuous strong lateral spine (sg 1) on each side; dorsally, the segment presents a pair of rounded evaginations, more or less prominent, confluent at the midline, which are associated with dorsal warty sensory areas; sometimes an additional pair of conical dorsolateral evaginations is found. Thoracomer 13 (second genital segment) with a strong ventrolateral spine (sg 2) on each side, just above the brood pouch (Figs. 10 F, G, H); dorsally, this segment presents a pair of usually prominent sensory areas. Abdominal segment II bearing a pair of ovoid ventrolateral denticulate protuberances (vp) (Fig. 10 E) as in male, showing diverse degrees of separation from each other in different specimens (Fig. 7 G, H). These protuberances may be hidden by the brood pouch (Fig. 10 F). Similar but smaller structures were also observed in abdominal segments IV and VI. Ovaries T-shaped, extending up to abdominal segments I – IV, or more rarely V. Brood pouch subcylindrical and rounded at the tip, without ventrolateral outgrowths, extending to abdominal segments IV – V (Fig. 10 G, H); however, in Bo- 18 it is particularly longer, reaching abdominal segment VI or even slightly surpassing the abdomen length (Fig. 10 F). Uropods (or cercopods) length: 1.2 mm (0.8 – 2.2; n = 30), markedly shorter than in male (Tables 3, 4); on average 2.4 times as long as the anal segment (or telson) (1.9 – 3.4; n = 30) (Table 4), divergent, uniformly provided with plumose marginal setae all over. Cysts (resistant eggs). Only type I cysts were found in the brood pouches of the new species. Cysts spherical (Fig. 11 A), mean diameter: 223 µm (187 – 242 μm; n = 7). Surface sculptured with ridges defining numerous cells (9 to 11) across the egg diameter, irregularly sized, subcircular or oval; mean dimensions: 28 – 35 µm (20 – 44 to 33 – 45 µm; n = 22), cells either isolated or coalescent (more frequently in pairs up to 5 cells that are delimited by narrower and lower ridges), cell groups either lined up or forming diverse arrays; each cell forms a shallow depression with grained bottom (Fig. 11 C); ridges are steep-sided and narrow except at intersections. Internal alveolar layer made of thin trabeculae (0.8 – 1.0 μm) separating polygonal or spherical vesicles that gradually decrease in size from the cortical (0.5 – 0.6 μm) to the basal zone (0.3 – 0.4 μm), where they are densely packed (Fig. 1 1 E). Size. Male total length from the front to the tip of uropods (or cercopods) excluding setae: 14 mm (10 – 22 mm; n = 26); measurements of other characters in Table 3. Female total length from the front to the tip of uropods (or cercopods) excluding setae: 14 mm (11 – 22 mm; n = 30); measurements of other characters in Table 4. Differential diagnosis. The morphological diagnostic characters of the two species of the genus Archaebranchinecta from the South American Altiplano are summarized in Table 5.
A new species of Archaebranchinecta (Anostraca: Branchinectidae) from the South American Altiplano