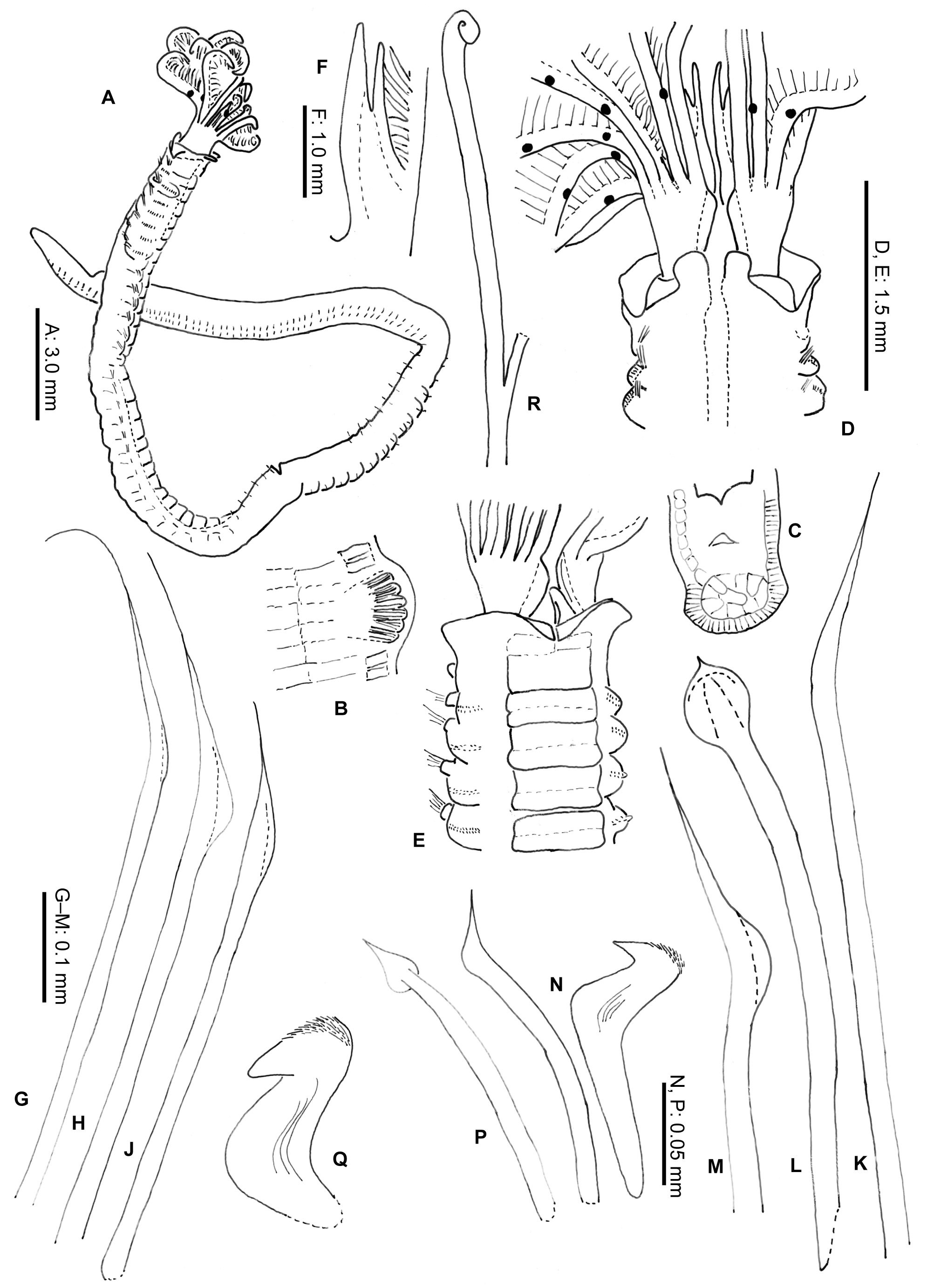
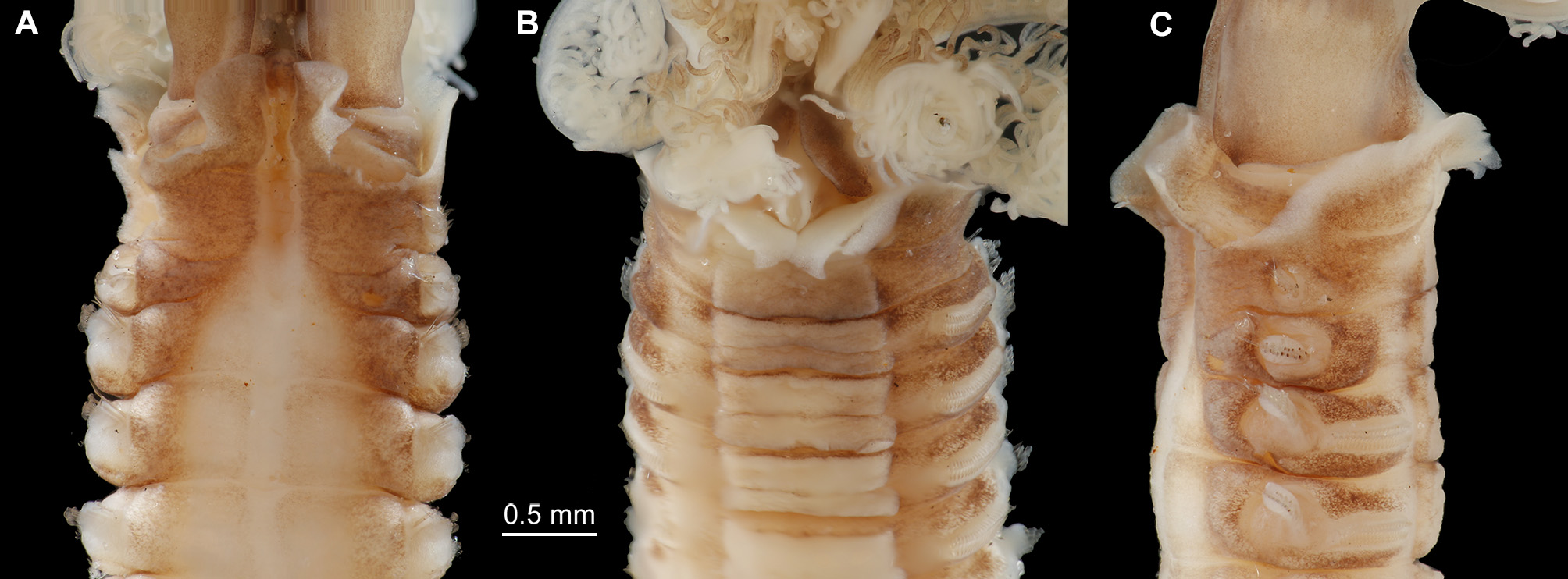
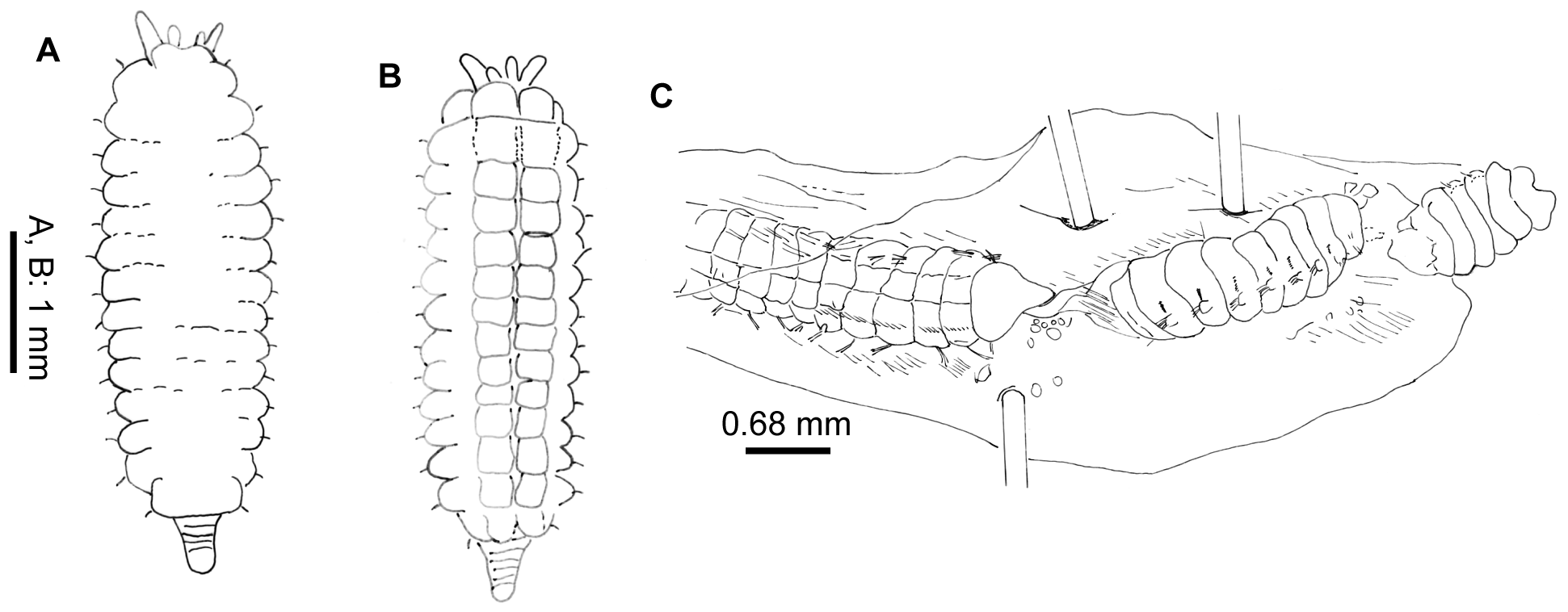
Description. Neotype complete in two parts. Body without radiolar crown about 38 mm long, 1.5 mm wide. Radiolar crown 4 mm long. Thorax with 8 segments and abdomen with about 120 (Fig. 1 A). Radiolar crown asymmetrical with radioles (24) shorter ventrally, tips short and blunt. Dorso-lateral radioles with one or two (can be up to four) unpaired, dark compound eyes, except the dorsal most pair (Fig. 1 B; R x 2121 xx 11 xxx, L x 11221211 xxx). Interradiolar web absent. Radiolar lobes about 7.5 mm long, with dorsal and ventral basal flanges (Fig. 1 D), ventral ones more oblique to accommodate ventral sacs (Fig. 1 E). Long dorsal lips about half length of radioles with a bifid appearance, each being supported by both radiolar appendage (mid-rib) and enlarged pinnule from base of adjacent radiole (Fig. 1 F). Dorsal collar (Fig. 2 A) with two high, rounded lappets extending wellabove junction of crown and thorax; mid-dorsal margins of dorsal lappets fused to sides of midline faecal groove; dorsal collar lappets with lateral margins in line with body axis anteriorly and oblique posteriorly, part of a wide, Vshaped notch showing peristomium (Fig. 1 D). Lateral collar (Fig. 2 C) oblique anteriorly, highest ventral part close to midline incision and in front of ventral sacs. First ventral shield (Fig. 2 B) twice as long as following ones, anterior margin indented medially, following thoracic shields rectangular, each divided transversely, but indistinctly, into two halves, grooves separating shields well defined (Fig. 1 E). Collar chaetae with two rows of broadly hooded chaetae, hoods as wide as or narrower than handles (Figs 1 G-H, J). Superior notochaetae of following thoracic fascicles narrowly hooded (Fig. 1 K), few, slender and longer than inferior chaetae, each inferior paleate chaeta (about ten) with small distal mucro (Fig. 1 L). Abdominal neurochaetae of one kind, elongate broadly hooded, each with hood twice width of handle (Fig. 1 M). Thoracic tori short with gaps between ventral ends and lateral margins of shields (Fig. 1 E), each uncinus avicular, with numerous fine crest teeth covering half of main fang, distance between end of handle and breast nearly twice as long as that between breast and tip of main fang, each companion chaeta with longer handle than that of uncini (Fig. 1 N) and distal angled, tear-drop blade (Fig. 1 P). Abdominal uncini smaller than thoracic and with shorter handles (Fig. 1 Q). Pygidium bilobed. Tube, enrolled distally (Fig. 1 R), of thin semi-hardened mucus with outer layer of embedded detritus including fragments of black and white shell in emergent areas, sometimes found with posterior branch made by departed offspring. Colour. Live specimens with liver-coloured pigments in broad bands around radiolar crown, on outside of crown base, on collar, surrounding 1 st and 2 nd parapodia and forming oval patches each side of 3 rd and 4 th parapodia. Paler tint flanking dorsal faecal groove and covering 2 nd and 3 rd ventral shields; rest of body pale and most ventral shields paler. Fixed specimens pale. Variation. The observed variability is correlated to the presence of asexual reproduction. A large clump of 34 specimens of Pseudopotamilla reniformis from Hvalfjordur (collected 8.10.1994) was examined to see the frequency of scissiparity. One tube contained an ‘ adult’, thorax and abdomen 37 mm long with a recently healed posterior (no pygidium) and a developing radiolar crown (two semi-circles of finger-like proto-radioles). Posterior to the parent were seven fragments of abdomen (largest 5 mm long) all with buds at both ends, only one with a distinct pygidium. One of these, a), with 7 segments, is 8 mm long, 3 mm wide, with crown an additional 1.5 mm long. The first and second segments have new parapodia with very small tori on the ventral sides of fascicles; segments 3 – 6 have both new thoracic and old abdominal parapodia, the latter with tori on the dorsal sides of fascicles. After fission, some original abdominal parapodia degenerate (Gross & Huxley 1935; Kolbasova et al. 2014), as new ones develop fascicles and tori arranged as in the thorax. The new complement of thoracic chaetigers will often be variable in number. The remaining abdominal segments retain their parapodia, the most posterior segment producing a new pygidium and new abdominal segments anterior to the pygidium. Another fragment, b), has a body 5 mm long, 1.7 mm wide, with a developing thorax of seven segments each with both new and old parapodia. The abdomen is left with nine original segments followed by new, short, narrow segments anterior to a new pygidium (Fig. 3 A – B). Many of the other thirty-three parents also lacked fully developed crowns. If the radioles were well developed, the pinnules were often under-developed and only three had compound eyes. Five specimens have eight thoracic segments, five specimens have nine segments, sixteen have ten segments, and five have eleven segments. These specimens produced 19 scissiparous offspring between them, ten with buds at one end, nine with buds at both ends and only two with a developing crown and posterior abdomen. Fourteen collections of Pseudopotamilla reniformis from Nova Scotia and Newfoundland showed patchy distribution, but at stations 1, 2 and 9 (July and October, see Materials and Methods) they were found in considerable aggregations typical of asexual reproduction. The contents of nine tubes from station 2, Digby Neck (largest 69 mm long and 2 mm wide and the smallest 8 mm long and 0.7 mm wide) were examined (MEP). Four of the smaller tubes (8 to 50 mm long) had each branched from a larger parent tube, 25 mm or more from the tube aperture; two specimens in these tubes (now labelled 5 and 7) contained nearly entire small adults, but only one with a radiolar ocellus. One is 17 mm long with about 65 chaetigers and the other 11.3 mm long with 24. The former lacked offspring, but the latter had two. The largest adult in these four tubes was 23.9 mm long, with 28 chaetigers and a nearly complete crown (radioles without ocelli). It had dehisced posteriorly to produce three abdominal fragments (Fig. 3 C): fragment a) 4 mm long with 12 segments and developing radioles; fragment b) 2.5 mm long, 9 chaetigers and no radiolar buds and fragment c) 1.4 mm with 5.5 chaetigers, also without radiolar buds. Of the adults in the five larger non-branching tubes, four had radiolar crowns and three had 1 – 2 dehisced abdominal fragments. Only one out of the ten fragments examined showed any sign of re-organisation of fascicles and tori, from the abdominal to the thoracic arrangement. The only noticeable difference between samples collected in July and August from eastern Canada and in June from Iceland is that thirty-one of thirty-four Icelandic adult specimens had re-developing radiolar crowns. Crowns can be lost by predation, but it seems unlikely that so many adults within the Hvalfjordur clumps could have lost their crowns in such a way, as their speed of withdrawal into tubes is so very fast. Such parents were fairly large (maximum body width 3 mm) so it is a puzzle as to why the development of their crowns had been delayed. This contrasts with material collected from Sandgerði in July 2001, most of which had fully formed crowns. Six of the nine specimens from Nova Scotia also had crowns. The uniformity of development of scissiparous offspring in all three collections and particularly the sparseness of chaetal re-organisation in fragments examined, suggests their asexual development had been more or less synchronous.
Knight-Jones, Phyllis, Darbyshire, Teresa, Petersen, Mary E., Tovar-Hernández, María Ana (2017): What is Pseudopotamilla reniformis (Sabellidae)? Comparisons of populations from Britain, Iceland and Canada with comments on Eudistylia and Schizobranchia. Zootaxa 4254 (2): 201-220, DOI: 10.11646/zootaxa.4254.2.3