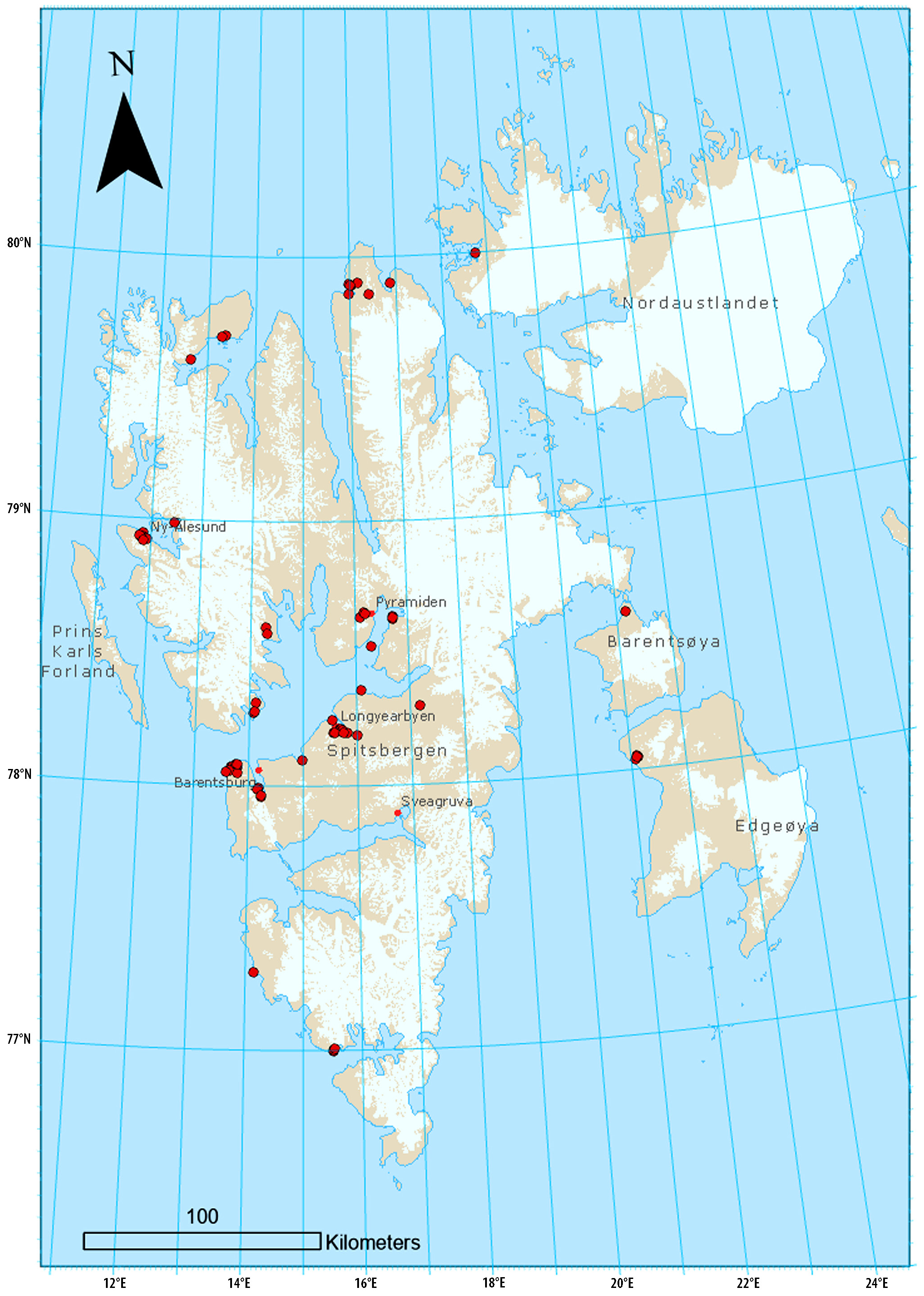
DISPERSAL Large Branchiopoda possess a remarkable ability to colonise new areas due to their dormant eggs within the egg banks (Longhurst 1955). Numerous studies have shown that branchiopod eggs are passively dispersed by mammals, birds, fish, amphibians, crayfish, insects, and wind (see Rogers 2014). When eggs are moved by predators like ducks and waders, which select similar habitats, this constitutes a ‘ directed’ passive dispersal vector. Additionally, eggs, larvae, and adults drifting downstream can also be considered ‘ directed’ passive dispersal, while wind acts as a ‘ random’ passive dispersal vector. Many branchiopods have developed the tactic of attaching their propagules to firm substrates, effectively reducing the likelihood of dispersal (Fryer 1996). This is also true in L. arcticus (Lakka 2013). This indicates that remaining in a favourable habitat may be more advantageous than risking the hazards of dispersal to other locations. An example of this is the slow colonisation of L. arcticus to newly established habitats near Aldegondabreen in Spitsbergen (Walseng et al. 2018). In Svalbard, small ponds are closely linked to the nearby terrestrial landscape, which often is inundated by snowmelt and rainfall, with passive water dispersal being possible (Borgstrøm et al. 2018). In the Scandes, the drift of larvae from upstream lakes into a reservoir is well documented (Brabrand et al. 2015). Downstream dispersal of adults of L. arcticus from a fishless headwater pond has similarly been observed in the Tornionjoki catchment in northern Finland (Lakka, unpubl.). Consequently, the establishment of L. arcticus in reservoirs downstream from their natural lower limit may also result from passive drift from upstream lakes (Dahl 1932; Aass 1969). Ponds and wetlands in Svalbard and lakes in the Scandes serve as important habitats for a diverse range of waterfowl and other bird species. At least 23 bird species are identified as potential dispersal vectors since they are frequently observed on or near water bodies (Lakka 2013, 2020; Håland 2012). Many of these birds prey on the adults, larvae and eggs of L. arcticus, thus serving as dispersal vectors. Water birds are more likely to travel directly between suitable habitats. Branchiopod eggs resist avian digestive enzymes, and passage through the alimentary canal can even enhance hatching (see Rogers 2014). Sticky eggs can be embedded in mud and attached to the fur of mammals or the feathers of birds (Neretina et al. 2020; Rogers et al. 2021). In Svalbard, eggs have been found at the foot of a ringed plover, Charadrius hiaticula Linneus, 1758 (see Lakka 2013). The distribution of similar haplotypes of L. arcticus across wide geographical ranges suggests high dispersal via migrating birds (Hessen et al. 2004). Lepidurus arcticus may also be actively dispersed by human stockings (Berg 1954; Aass 1969; Hesthagen 1979; Fjellheim et al. 2007). PREY-PREDATION RELATIONSHIPS Freshwater organisms play important ecosystem roles in linking terrestrial and aquatic detritus and periphytic algal production to higher trophic organisms such as fish. As an adult, L. arcticus is an omnivore species preying on detritus, bacteria, and different kinds of plants and algae, and it is also a predator of other crustacean species (Jeppesen et al. 2001), including conspecifics (Miller 1980; Lakka 2013, 2015). Evidence from a study of ten lakes on Hardangervidda conducted between 2000 and 2002, where the food web of brown trout was analysed using stable isotopes, revealed elevated δ 15 N levels indicating L. arcticus’ diverse diet in comparison with other invertebrates (Rognerud et al. 2003). In Arctic freshwater communities, L. arcticus dominates in size and biomass (Klemetsen et al. 1985; Dimante-Deimantovica et al. 2015, 2018). As a predator of smaller crustaceans, it significantly impacts the planktonic food web, as demonstrated in a study of Greenland ponds and lakes (Cristoffersen 2001). However, research on 75 ponds and lakes in Svalbard showed no substantial difference in taxa numbers between locations with and without L. arcticus (Walseng et al. 2018). Many lakes in the Scandes are renowned for their excellent fishing conditions, hosting large, high-quality brown trout (Figure 7). The abundance of crustacean food sources like L. arcticus, Gammarus lacustris G. O. Sars, 1863, and Eurycercus lamellatus (O. F. Müller, 1776) contributes to the brown trout’s substantial growth, often reaching one to three kilos or more (Qvenild et al. 2021). Arctic charr also attain big-sized specimens of excellent quality in lakes with these crustaceans. Lepidurus arcticus is highly sought for by fish. When this species is abundant, it positively influences the growth and production of the fish stocks, which are of considerable economic importance to Northern countries. In Lake Øvre Heimdalsvatn, L. arcticus accounted for 15 % of the annual energy intake of the brown trout population (Lien 1978). Instances of heavy predation show more than 200 – 300 specimens in a single brown trout stomach (Aass 1969; Borgstrøm 1970). Lake Sylvetjønne, located at high altitude in the Jotunheimen mountains, demonstrated remarkable yields of brown trout, primarily based on a diet of L. arcticus and G. lacustris, with total yields reaching 13 kg per hectare in 1941 – 1942 and individual specimens weighing up to 10 – 12 kg (Hesthagen 2005). Brown trout selectively prey on L. arcticus, with its significance as a food source increasing with the size of the fish (Qvenild & Hesthagen 2020). Conversely, an overstocking of juvenile brown trout can negatively affect L. arcticus populations. For example, in Lake Vesle Øljusjø, excessive stocking led to the species’ nearextinction, although it reappeared in brown trout diets after a decade without stocking (Aass 1969). In a comprehensive dietary analysis of brown trout populations across five lakes on Hardangervidda, the exceptionally strong 1997 year-class nearly eliminated L. arcticus and G. lacustris (Qvenild & Hesthagen 2020). The crustacean populations recovered once the brown trout numbers returned to normal levels. The absence of L. arcticus in High Arctic lakes is frequently attributed to the presence of Arctic charr, which is the only fish species present (Jeppesen et al. 2001; Presthus Heggen et al. 2010; Borgstrøm et al. 2018). In the Scandes, the invasive Eurasian minnow has populated many lakes and reservoirs since the 1960 s, thriving in habitats up to 1400 m a. s. l (Museth et al. 2007). As opportunistic foragers, minnows introduced to high-altitude lakes have been linked to reduced availability of brown trout prey (Museth et al. 2007; Borgstrøm et al. 2010). In Lake Øvre Heimdalsvatn, minnows appeared in 1969, leading to significant changes in the brown trout diet during the summer months between 1970 – 1972 and 1993 – 2005 (Lien 1978; Borgstrøm et al. 2010). Here, the Eurasian minnow has severely reduced L. arcticus populations, causing its virtual disappearance from the brown trout diet. On Hardangervidda, the Eurasian minnow was first observed in Lake Ørteren in 1973 and has since spread to at least 42 lakes in the northeastern plateau (Qvenild & Hesthagen 2019). The expansion continued westward, eventually reaching the headwater lakes in the tributary Sevra in the Lågen catchment. Numerous lakes in the neighbouring catchments, where both L. arcticus and G. lucustris are staple food organisms for brown trout, are easily accessible from the flat terrain near Stigstuv. In August 1999, a rotenone treatment was carried out, followed by another in September 2000, effectively eradicating the minnows. This action also caused immediate mortality within the invertebrate community (Fjellheim 2004). Remarkably, the diversity was almost completely restored within just one year. Lake Skjerja, formerly a prominent brown trout lake, experienced drastic declines in brown trout yield due to intense exploitation and subsequent minnow invasion, leading to a reduction in annual catches to only 32 % of the initial values (Qvenild et al. 2024). Despite fishing efforts to control minnows, the competition for food has negatively impacted brown trout production. In Lake Skjerja, the two crustacean species, G. lacustris and L. arcticus, were the basic food items. Fishing with baited traps on the minnows and the presence of a large fraction of predatory brown trout feeding on minnows have reduced the predation pressure on these two crustacean species, and they are still a part of the brown trout diet. In contrast, L. arcticus and G. lacustris no longer seem to be important in the brown trout diet in the nearby Lake Skaupsjøen, where no systematic reduction of Eurasian minnow is performed (Borgstrøm 2009). The balance of fish predation on L. arcticus is mainly determined by the abundance of fish and the availability of suitable refuges for the prey. Unlike other notostracan species that grow rapidly in temporary water bodies before they dry out or encounter increased predation (Longhurst 1955), this strategy is unfeasible in the High Arctic’s permanent lakes, which are often inhabited by stunted populations of Arctic charr dwarfs (Jeppesen et al. 2001). In these cold environments with clear water, the growth and development of L. arcticus are slow, making them particularly vulnerable to fish predation. Fish density also acts as a limiting factor in the permanent lakes of Bear Island and the Scandes. Nevertheless, in lakes where optimal refuges are accessible and fish abundance is low, L. arcticus can thrive. Thus, L. arcticus’ adaptability to various habitats seems to be broader compared to other notostracan species. Studies conducted on Hardangervidda and Northern Finland indicated a higher occurrence rate of L. arcticus populations in larger lakes with better access to refuges (Qvenild & Hesthagen 2019; Lakka 2021). Larger lakes also are generally colder than smaller ones. Birds are significant predators of L. arcticus. In Spitsbergen and Bear Island, species such as the purple sandpiper Calidris maritima Brünnich, 1764, the dunlin Calidris alpine (Linneus, 1758), and the arctic tern Sterna paradisaea Pontoppidan, 1763, frequently prey on L. arcticus in the shallow tundra ponds (Summerhayes & Elton 1923; Lakka 2015). When faced with these avian predators, L. arcticus exhibit behavioural adaptations (Lakka 2013). Observations in a pond with a dense L. arcticus population in Kapp Linné in Svalbard revealed that L. arcticus sought the shallowest areas when a red phalarope, Phalaropus fulicarius (Linneus, 1758), visited. The shallow and muddy waters (5 – 15 cm deep) hindered the red phalarope from performing its typical hunting behaviour of swimming in circles. The variations in the colour morphs of L. arcticus are believed to be an adaptation to avoid bird predation in addition to harmful UV radiation (Lakka 2013). In Lake Langavatnet on Hardangervidda, sea ducks, such as the black scoter Melanitta n. nigra (Linneus, 1758), have declined comparatively with the increase in brown trout abundance, which likely is a result of increased competition for their common food item, L. arcticus (Håland 2012). Simultaneously, piscivorous waterbird species like the Goosander (Mergus merganser Linnaeus, 1758), Red-breasted Merganser (Mergus serrator Linnaeus, 1758), Great Cormorant (Phalacrocorax carbo Linnaeus, 1758) and Black-throated Diver (Gavia arctica Linnaeus, 1758) grazing on the enhanced numbers of small brown trout increased. ENVIRONMENTAL CONDITIONS Lepidurus arcticus has demonstrated a remarkable ability to withstand significant environmental changes in the harsh alpine and Arctic environments since its emergence. Despite this, the species exhibits considerable sensitivity at various stages of its life cycle to numerous environmental stressors. Critical temperatures and thermal thresholds Temperature is a key environmental factor controlling the distribution of ectotherm species, including branchiopods. Typically, Triops species are more warmth demanding than Lepidurus (Fryer 1988). The cold-adapted L. arcticus can withstand water temperatures ranging from 1.8 ° C (Lakka 2013) to 19 ° C (Arnold 1966). Although L. arcticus can endure temperatures above 20 ° C for brief periods (Borgstrøm 2019), it is crucial that temperatures do not remain above 16.5 ° C (Lakka 2020). The ideal temperature for adult L. arcticus is around 10 ° C (Lakka 2013). In the Arctic, this optimal thermal condition is met for only short-duration periods, but it is more commonly encountered in Scandes lakes. Every species has an optimal temperature range for hatching efficiency. In a laboratory experiment in Svalbard, the highest hatching rate for L. arcticus was noted at 10 ° C (Pasquali et al. 2019). There was no hatching observed at 15 ° C and 25 ° C, likely because these temperatures surpassed the eggs’ physiological tolerance limits. Comparatively, the optimal hatching temperature of the more southerly distributed L. couessi was found to be 20 ° C (Hann & Loonsberry 1991). On the other hand, the more adaptable species, T. cancriformis, exhibited hatching within a broader range of 16 – 32 ° C, with an optimal range of 22 – 32 ° C (Kuller & Gasith 1996). Elevated temperatures above 15 ° C during the juvenile phase negatively impact survival (Pasquali et al. 2019). Abrupt changes in water temperature during the initial hatching phase, following ice break-up, can frequently occur. For instance, Lake Øvre Heimdalsvatn experienced a rise in water temperature from 4 to 14 ° C within just ten days in 2018 (Sildre, https: // sildre. nve. no). These temperature shocks are especially likely in small, shallow lakes and ponds, particularly in the Scandes region. During the moult stage, L. arcticus is also highly vulnerable (Thiéry 1997). Warm, oxygen-poor water can trigger an ‘ oxygen search behaviour’, where an adult swims to the surface, collects a small air bubble between their legs, and then swims back to the bottom (Lakka 2013). This behaviour exposes L. arcticus to predators such as birds and fish. The lower thermal threshold The time required for an organism to complete its life cycle can be measured in thermal units, known as degree-days. Laboratory experiments conducted with L. arcticus eggs collected in the autumn from lakes in southern Norway (Borgstrøm & Larsson 1974) and northern Finland (Lakka, unpubl.) suggest that a cumulative thermal energy of between 400 and 500 degree-days is necessary for the embryonic development to complete once the eggs are submerged in water. Fryer (1988) also noted that L. arcticus eggs could hatch without undergoing a drying or freezing period. In dry or frozen conditions, embryonic development is halted. For example, the shallow Lake Solvatnet in Spitsbergen freezes solid during winter (Pasquali et al. 2019). Eggs collected after the ice melted and incubated at temperatures of 5.0 ° C and 10.0 ° C began hatching after 15 and 7 days, respectively. Thus, eggs incubated at 5.0 ° C in Lake Solvatnet hatched more quickly than those kept constantly wet (Borgstrøm & Larsson 1974). This indicates that part of the embryonic development must have occurred during the previous summer or earlier. Only 64 % of the eggs in the Lake Solvatnet experiment hatched, highlighting the importance of the egg bank as a buffer against variable environmental conditions (Hann & Lonsberry 1991; Brendonck 1996). Lepidurus arcticus can tolerate very low temperatures close to freezing. However, a minimum thermal input is essential for the organism to develop into adults. In a study of six lakes on Hardangervidda, no adult L. arcticus was found in brown trout stomachs until approximately 400 degree-days were achieved (Qvenild et al. 2018). This equates to a mean summer temperature of at least 5 ° C from July 1 st to September 15 th. In the High Arctic, only ponds tend to be warm enough to support L. arcticus (Nowiński & Wiśniewska-Wojtasik 2006; Lakka 2013) in at least some seasons, enabling the establishment of an egg bank. After hatching, the postembryonic phase must be completed, and new eggs laid before freezing. In particularly cold environments, females seem unable to produce eggs (Olofsson 1918; Lakka 2015), thus restricting their altitudinal and latitudinal range. The heat sum required may vary in different climatic regions since adaptation to colder habitats is likely, as is revealed in Arctic charr (Koops & Tallman 2004; Jeuthe et al. 2016). However, information is lacking for L. arcticus. Late ice break-up and low water temperatures during a short growing season can significantly impede the growth and development of L. arcticus. This has been documented over several years on Hardangervidda (Halvorsen 1973; Pedersen & Scobie 1990; Simonsen & Valderhaug 1994; Borgstrøm 2016; Qvenild et al. 2018) and in ponds in Svalbard (Lakka 2013). Lepidurus arcticus was found to be smaller in colder ponds near glaciers compared to warmer ponds closer to the sea (Lakka 2013). In many ponds studied by Olofsson (1918), L. arcticus appeared to be absent, likely due to excessively low temperatures. The upper thermal threshold The lower altitude boundary for lakes containing L. arcticus in the Scandes signifies that beyond a certain thermal threshold, the conditions become unfavourable. Most Lepidurus lakes remain at mean air temperatures below the 10 ° C isotherm (Qvenild et al. 2021). This explains why the distribution appears to be dichotomous in the Scandes, splitting into northern and southern zones. The intermediate region is generally too warm to support L. arcticus populations. In the northernmost area, reaching up to 70 ° N, the 10 ° C isotherm falls below 250 m a. s. l, with few lakes situated above 300 metres in this region. Consequently, the likelihood of discovering new L. arcticus populations is minimal. Few Lepidurus lakes have a mean lake air temperature higher than 11 ° C in summer (Qvenild et al. 2021). Lakes with lake air temperatures at this level tend to have mean summer water temperatures close to 14 ° C. Hence, the upper thermal threshold for Lepidurus lakes was assumed to be close to this level. Excessive accumulation of thermal energy during the initial embryonic phase can lead to life cycle mismatches, resulting in eggs hatching too early under completely dark conditions of winter. In the Scandes, L. arcticus is rarely found in ponds, although it does occur (Berg 1954; Koli 1957; Halvorsen 1973; Hesthagen 1979; Blomkvist 1995; Walseng et al. 1996). Most small lakes and ponds have environmental conditions that are often too unstable for sustained survival. Fluctuating temperatures and acidic waters have adversely impacted these habitats (Blomkvist 1995; Fjellheim et al. 2002; Lakka 2013, 2020). Lepidurus arcticus populations are more commonly found in larger lakes where there is better access to refuges and colder water (Qvenild & Hesthagen 2019; Lakka 2021). Moreover, deeper lakes allow L. arcticus to evade warm surface layers by migrating to cooler depths. Further north, including Bear Island and Spitsbergen, the species predominantly inhabits coastal zones (Klemetsen et al. 1985; Lakka 2013). In these northern islands, the mean air temperature during summer typically does not exceed 5 ° C, ensuring that elevated temperatures do not limit its distribution. Water quality Acid waters low in calcium In addition to temperature, water quality plays a crucial role in determining distribution. These conditions are likely to interact with one another, as evidenced in T. cancriformis (Schönbrunner & Eder 2006). In a study on Hardangervidda, the absence of L. arcticus in the western region was attributed to low water temperature, a short growing season, and water that was low in pH and calcium (Qvenild & Hesthagen 2019). The melting snow in spring usually leads to a drop in pH levels during ice break-up in Arctic and alpine lakes (Fjellheim et al. 2002; Lakka 2020), which can be detrimental or even lethal for L. arcticus larvae, as shown in a laboratory study by Borgstrøm & Hendrey (1976). Hence, the juvenile phase is crucial in evaluating extinction risks. For crustaceans, rapid calcification of their exoskeleton immediately after moulting is essential, but this process can be hampered in regions with low calcium levels and poor water quality (Rukke 2002). This problem is further exacerbated by low pH levels, which normally interfere with low calcium concentrations. Multiple studies have shown that L. arcticus does not inhabit water bodies with pH levels below 6.1 (Borgstrøm & Larsson 1974; Borgstrøm & Hendrey 1976; Fjellheim et al. 2007; Lakka 2013). However, a recent analysis of 95 lakes on Hardangervidda, which host L. arcticus, revealed that pH levels ranged from 5.49 to 7.26, and calcium levels varied between 0.28 and 5.54 mg / L - 1 (Qvenild & Hesthagen 2019). Ten of these lakes had pH levels under 6.0 and calcium levels below 0.84 mg / L - 1, conditions which are thought to be critical for L. arcticus. The two lakes with the lowest calcium levels, Lake Svartavasstjørni and Svartavatnet, have been monitored each year. In 1993 – 1994, the pH was approximately 5.5, and the calcium level was approximately 0.4 mg / L - 1 (Tysse et al. 2022). An annual liming programme began in 1994, and the lost L. arcticus population was restocked over three consecutive years (1997 – 1999) (Fjellheim et al. 2007). The species has been observed nearly every year since then, despite the liming programme ending in 2013. Calcium levels have stabilised between 0.5 and 1.0 mg / L - 1, and pH values have been close to 6.0. Therefore, L. arcticus can survive in waters with calcium levels at approximately 0.5 mg / L - 1 and low pH values near 6.0. Proximity to refugia with better water quality is crucial for mitigating harmful pH fluctuations (see Fjellheim et al. 2007), as close to non-lethal conditions may inadvertently impact functional traits like growth and reproduction. Previous studies have indicated that L. arcticus likely became extinct in multiple regions, such as Finland (see Lakka 2020) and Northern Sweden (Lingdell & Engblom 2002). Recent research on the ponds examined by Koli (1957) suggests that the population has become extinct due to acidification (Lakka 2020). In the southernmost regions of the Scandes, specifically Setesdalsheiene, numerous populations of L. arcticus may have similarly faced decline and eventual extinction following the disappearance of brown trout populations observed in the mid- 1880 s (Enge et al. 2017). Humic waters The concentration of total organic carbon (TOC mg L - 1) in humic waters is an indicator of the runoff of external organic matter from surrounding areas. In shallow inland water bodies, carbon can accumulate at the bed of lakes or ponds, influencing the carbon flux. For lakes with clear water in the Scandes, TOC levels are typically low. On Hardangervidda, the TOC in nearly all Lepidurus lakes was less than 2.0 mg C L - 1 (Qvenild & Hesthagen 2019). Research conducted by Lakka (2013) in Svalbard in 19 ponds revealed TOC levels ranging between 0.9 and 52.0 mg L - 1. Despite these conditions, thriving populations of L. arcticus were observed, demonstrating the species’ tolerance to elevated TOC concentrations, at least temporarily. Salinity The hatching of many notostracan species is influenced by various environmental factors, including salinity (Brendonck 1996; Schönbrunner & Eder 2006). The salinity levels in water bodies where L. arcticus resides are typically very low. Salinity can exceed 30 ‰ in coastal regions of the Barents Sea. Research from Spitsbergen has shown that L. arcticus flourishes in ponds with salinities reaching up to 1.5 ‰ (Lakka 2013). Slightly saline water has been found to promote the growth of L. arcticus, an adaptation likely linked to survival in temporary ponds where salinity increases as they dry up. In these environments, L. arcticus must efficiently produce eggs before the pond desiccates. This assertion is backed by two observations from Lakka (2013). Firstly, faster growth to a larger size enhances reproductive success in L. arcticus, as larger individuals can produce more and bigger eggs. A low salt concentration (1 ‰) was found to significantly accelerate the second moulting process. Secondly, smaller individuals began producing more eggs in slightly saline temporary ponds compared to permanent ones. Arctic lowlands near the sea and river systems are generally former seabeds where sea spray is prevalent. This is also true for Svalbard, where populations of L. arcticus are located near the sea (Lakka 2013). The salinity tolerance of L. arcticus may have evolved in coastal seabed habitats where the primary water source was melting snow and glacier ice, with permafrost keeping pond bottom soil activity minimal. Lepidurus arcticus in regulated lakes In the Scandes, L. arcticus is frequently observed in regulated lakes. Out of 483 locations where L. arcticus is present, 88 are hydropower reservoirs, of which 25 are regulated more than 20 m, sometimes even 125 m (Qvenild et al. 2021). These reservoirs present highly variable environmental conditions that are often detrimental to many freshwater organisms. However, L. arcticus and another Branchiopoda, the Cladocera Eurycercus lamellatus, typically show an increase in abundance (Dahl 1933; Huitfeldt-Kaas 1935; Aass 1969). This is likely due to their drought- and freeze-resistant eggs, which may stay dormant in the dry drawdown zone. The relative significance of these two species as food for brown trout was illustrated in the reservoir Lake Aursjøen in the Reinheimen area (Hesthagen 2018; Lakka et al. 2020). During the period 1985 – 1989, L. arcticus dominated in the brown trout diet. Following a change in the regulation regime, the abundance of L. arcticus collapsed. The reason remains largely unknown, but both climate- and hydropower-driven changes may be important. The alternative food source, E. lamellatus, did not appear to adequately compensate for the caloric deficit resulting from the decreased consumption of the larger L. arcticus. This likely contributed to the observed decline in the condition, growth, and delayed maturation of the brown trout. The temperature of the water significantly affects the altitudinal distribution of L. arcticus, also in reservoirs (Qvenild et al. 2021). This was initially demonstrated through the impoundment history of Lake Pålsbufjorden in the Lågen catchment (Dahl 1932). This catchment drains numerous Lepidurus lakes located in the eastern fells of Hardangervidda. Lepidurus arcticus was not previously recorded in this downstream lake, but just a few years after the impoundment, it became established in the new reservoir. Dahl (1932) explained this phenomenon by the cooling of the drawdown zone, leading to an ‘ arctification’ of the lake. Specifically, the ‘ arctification’ might be attributed to the halted embryo development in the dry drawdown zone, which accumulates fewer degree-days than in the wetter zone below. This allowed L. arcticus to flourish in Lake Pålsbufjorden, a habitat that was initially too warm for this species. Comparable altitudinal shifts of L. arcticus have been observed in other lakes following the establishment of hydropower reservoirs (Aass 1969; Brabrand & Saltveit 1980). The reproductive success of L. arcticus in regulated lakes is heavily influenced by the timing of reservoir refilling. In Lake Mårvatn on eastern Hardangervidda, increased lowering of the water level during the winter of 1969 / 70 without subsequent refilling resulted in poor condition and low brown trout catches (Borgstrøm 1973). In preceding years, L. arcticus was the main food source, but it almost disappeared from the diet in 1970 and 1971, likely because the eggs remained dormant in the dry drawdown zone. Moreover, siltation resulting from erosion adversely affected the production of algae, which serves as the primary food for L. arcticus larvae. During 2009 – 2010, Lake Mårvatn was again permanently lowered, resulting in a similar outcome (Rognerud & Fjeld 2014). Proper management practices are thus crucial to maintaining healthy populations of L. arcticus and supporting the ecosystems that depend on them. In some reservoirs, the introduction of new fish species has caused L. arcticus to become a less significant food source. For instance, in Lake Stolsvatn in the Hallingdal mountains, L. arcticus was a primary food item for brown trout before the Eurasian minnow was introduced (Borgstrøm et al. 1985). The eggs, which are reddish or orange, are typically deposited in the littoral zone, and the sticky eggs are firmly glued to the bottom substrate. Upon hatching, the larvae are highly visible and exposed to predation from the schooling minnows. After several years, adult L. arcticus no longer constituted a significant part of the brown trout diet. Minnows did not replace L. arcticus in the diet, indicating that new fish species can cause increased food competition and diminish brown trout yields. Similar adverse effects have been observed in other hydropower reservoirs such as Lake Halnefjorden on Hardangervidda (Lehmann et al. 2008) and Lake Kaldfjorden in the eastern fells of Jotunheimen (Hesthagen 2022). In Lake Kaldfjorden, Eurasian minnow and whitefish were introduced in the 1970 s, resulting in L. arcticus being excluded from the brown trout diet. The disappearance of L. arcticus in the fish diet may serve as an indicator of an impending decline in a fish population. In some reservoirs, the Eurasian minnow seems to be less harmfull. Lake Vinsteren in Jotunheimen was impounded in the 1940 s, and L. arcticus was the predominant food item for brown trout (Aass 1969). After the Eurasian minnow was introduced in the 1980 s, the abundance of L. arcticus has differed. However, there has been no significant decrease in brown trout yield, likely because the minnows are more exposed to brown trout predation, given the absence of refuges in the drawdown zone (Hesthagen & Gran 2019). CLIMATE CHANGE Few regions on Earth have experienced more dramatic climate changes than the High Arctic, a trend that is projected to continue unabated. Over the last thirty years, temperatures in Svalbard have increased by at least 2 ° C above the 1961 – 1990 normal (Hanssen-Bauer et al. 2019). Of particular interest in this context is the rise in summer temperatures. In the Svalbard archipelago, data from five meteorological stations reveal that the summer air temperature has increased by 1.41 ° C in the first two decades of this century compared to the normal period (July 1 st to September 15 th). Similar data from six meteorological stations in the Scandes show a concurrent increase of 1.26 ° C, suggesting significant changes in summer temperatures in both Arctic and Alpine regions. The Svalbard archipelago The diversity of freshwater organisms in the still waters of Svalbard is notably low, even when compared to other High Arctic regions (Coulson et al. 2014). Climate change is causing glaciers to retreat, leading to the formation of new, untouched freshwater ecosystems. This rapid warming and lengthening of growing seasons may influence biodiversity and dispersal patterns, potentially attracting more warmthdemanding species. Currently, there are 31 documented crustacean species on the Svalbard archipelago, with at least seven being recent arrivals (Dimante-Deimantovica et al. 2018). This represents an increase of over 20 % in the total number of known species on the island. As water bodies warm, L. arcticus may extend its range into the colder regions of Svalbard, including new inland ice-free areas, and become more prevalent on the colder islands, such as Nordaustlandet and other eastern islands. Dry sediments in temporary ponds are susceptible to wind dispersal. Increased bird activity due to warming affects ponds and lakes (Walseng et al. 2018), and birds preying on L. arcticus may aid in egg dispersal. The eggs can adhere to fur and are resistant to avian digestive enzymes (see Rogers 2014). Similar haplotypes over large areas suggest significant bird-mediated dispersal (Hessen et al. 2004). The distribution of freshwater fauna along a gradient from the glacier edge at Aldegondabreen in Grønfjorden to the coastline probably mirrors the colonisation history from the last 80 years of melting in Svalbard (Walseng et al. 2018). This study found L. arcticus in six of the 22 surveyed sites, all located in ‘ older’ water ecosystems near the shoreline (Bjørn Walseng, pers. comm.). So far, these findings suggest a relatively low dispersal rate of L. arcticus over a short period. Currently, L. arcticus exists below its optimal temperature in Spitsbergen and could thus benefit from warmer summer temperatures and prolonged growing seasons. Consequently, in the clear-water ponds in Spitsbergen, L. arcticus may experience accelerated growth and development. Notably, the size of L. arcticus has significantly increased since Sars’ studies in 1896 (Lakka 2013). The clear-water lakes may warm considerably and resemble those in Bear Island, potentially leading to coexistence with Arctic charr. Lakes such as Straumsjøen and Nordre Borgdam may reach temperatures conducive to developing such populations. The Scandes Sediment remains of L. arcticus indicate that it once had a much broader distribution across Europe (see Økland & Økland 2003), suggesting a later recolonisation of colder lakes formed its current presence in the Scandes. In the Scandes, L. arcticus mainly inhabits the alpine zone, with 87 % of its lakes located above the treeline (Qvenild et al. 2021). The early Holocene warm period (7800 – 7600 BP) serves as an early indicator of potential increased precipitation linked to ongoing greenhouse warming (Dahl & Nesje 1996). During this time, a notable rise in the treeline occurred, which could have resulted in a considerable range retraction if L. arcticus followed this treeline extension. Thus, prolonged warm periods might pose significant challenges for L. arcticus in the Scandes, which is the southernmost edge of its range in Europe. A retraction from the lower part of its distribution is documented for another cold-water adapted branchiopod, Branchinecta palludosa (O. F. Müller, 1788) (Lindholm et al. 2015). In the southern Scandes, L. arcticus extends its distribution slightly above 1500 m a. s. l. In a warmer climate, L. arcticus might potentially thrive some hundred metres higher than this. Baseline studies and ongoing monitoring programmes are urgently needed (Qvenild 2022). The only long-term programme in the Scandes, involving repeated monitoring of environmental conditions and biota, is at Lake Øvre Heimdalsvatn (Brittain et al. 2019). Since 1993, L. arcticus has virtually been absent from the brown trout diet in this lake. Predation from brown trout and Eurasian minnow poses a significant risk to L. arcticus, compounded by notable warming since 1985 (Sildre, https: // sildre. nve. no). The average summer temperature has nearly reached 14 ° C, a critical threshold (Qvenild et al. 2021). Consequently, further warming could be detrimental to L. arcticus in this lake. Updated surveys in Lepidurus lakes with observations older than ten years would be invaluable and might reveal extinct populations, particularly in the lowermost lakes.
Ecology and distribution of the Arctic Tadpole Shrimp Lepidurus arcticus (Branchiopoda, Notostraca) in the Fennoscandian Mountain Range and on the Svalbard Archipelago