Anthothela grandiflora
rosenknoppskorall(+8)·(Sars, 1856)
GBIF:2262167
0
Synonyms
PROFILE
Species Profile
Habitat
ABOUT
Descriptions(6)
Export occurrence data
Darwin Core Archive (ZIP)
GEOGRAPHY
Distribution Map
REGIONS
Geographic Distribution(36)
DATA
Occurrence Datasets
Common names used for this species across different languages and regions. Available in 3 languages and 1 country. 2 preferred.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide. This taxon has names in 3 languages. 2 names preferred.
CLASSIFICATION
Taxonomic Classification Tree

RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(27)
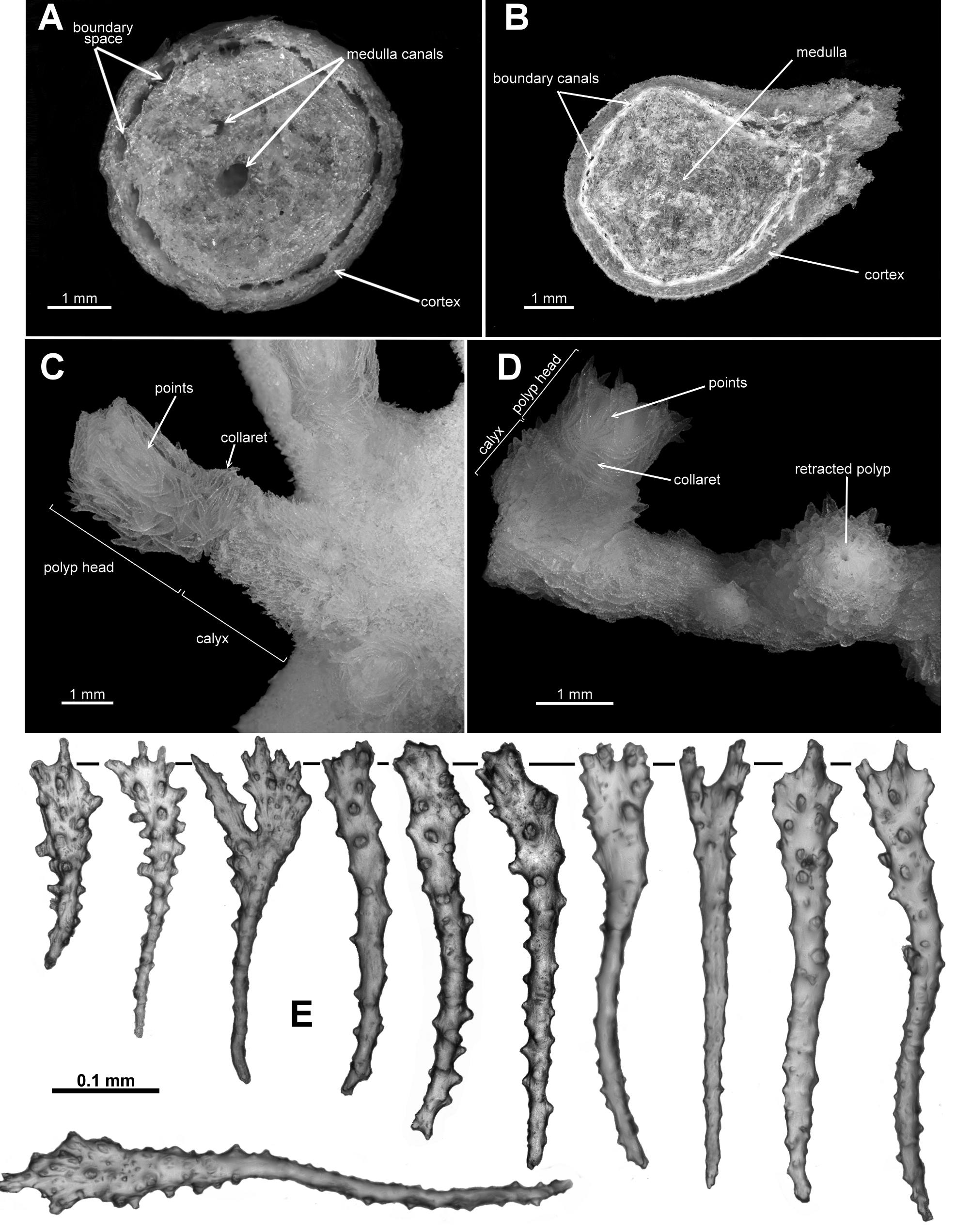
FIGURE 1. A. Cross-section of the holotype of Ƒictorgorgia eminens n. sp. showing extensive boundary canals which frequently anastomose and distinct central canals in the medulla; B. Cross-section of the holotype of Anthothela grandiflora showing boundary canals, adjacent but separate, and a medulla lacking any obvious central canals; C. Extended polyp of the holotype of A. aldersladei n. sp. showing polyp, calyx, points and collaret; D. Partly extended polyp of A. quattriniae n. sp. showing polyp head, calyx, points and collaret and, on right, a fully retracted polyp; E. Spatulate clubs found in Anthothela.

FIGURE 2. A. Josephinae clubs found in the pinnules and tentacle rachis of Ƒictorgorgia and Lateothela n. gen.; B. Spindles (93 – 95), needles (86 – 87) and bars (88 – 89) from Bayer et al. 1983, Plate 16; C. Sticks.

FIGURE 3. Sclerite forms found in the points, calyx and cortex of Anthothela and Ƒictorgorgia species.
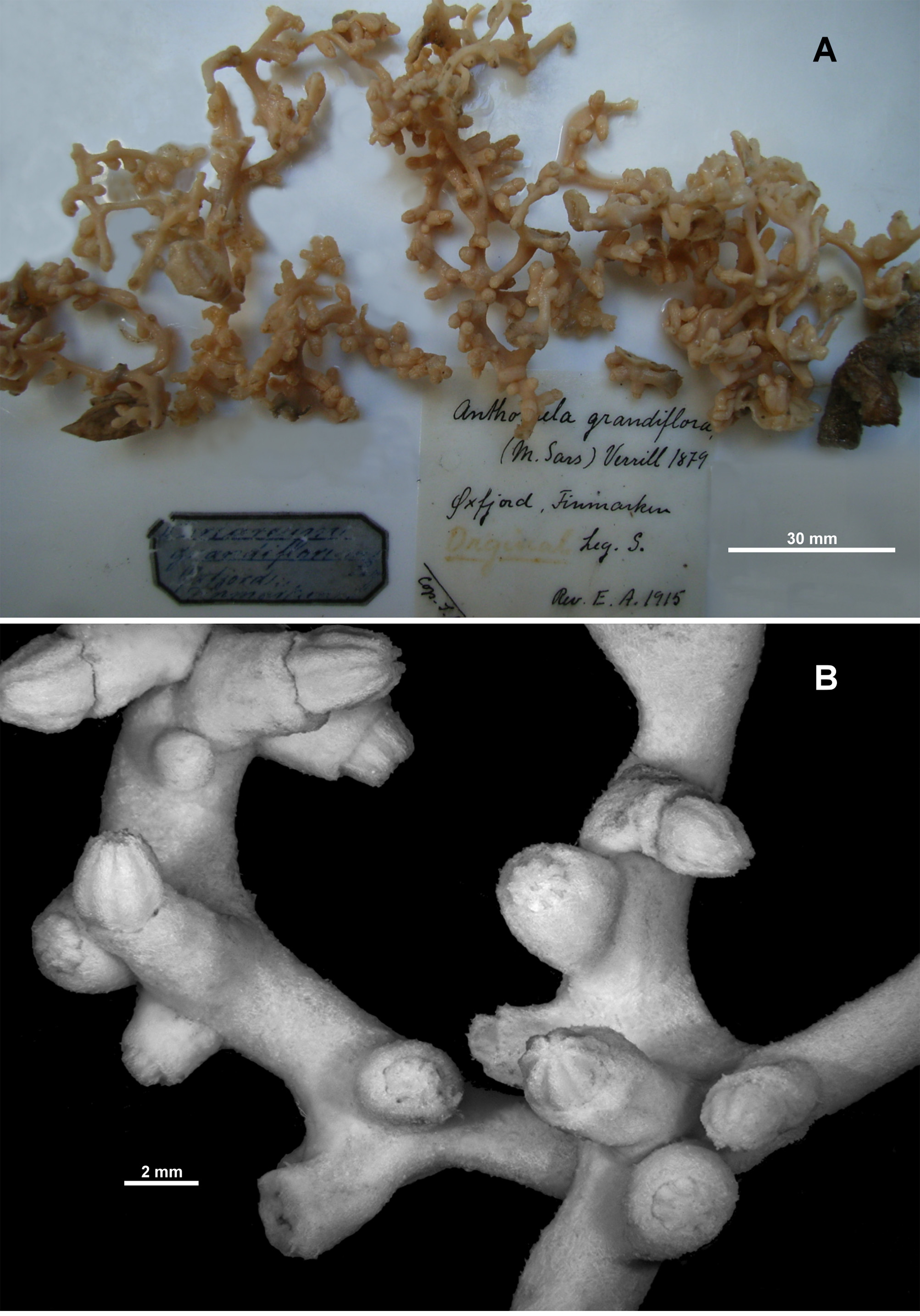
FIGURE 4. Anthothela grandiflora (Sars, 1856), holotype: A. Holotype fragments; B. Polyps and branches.
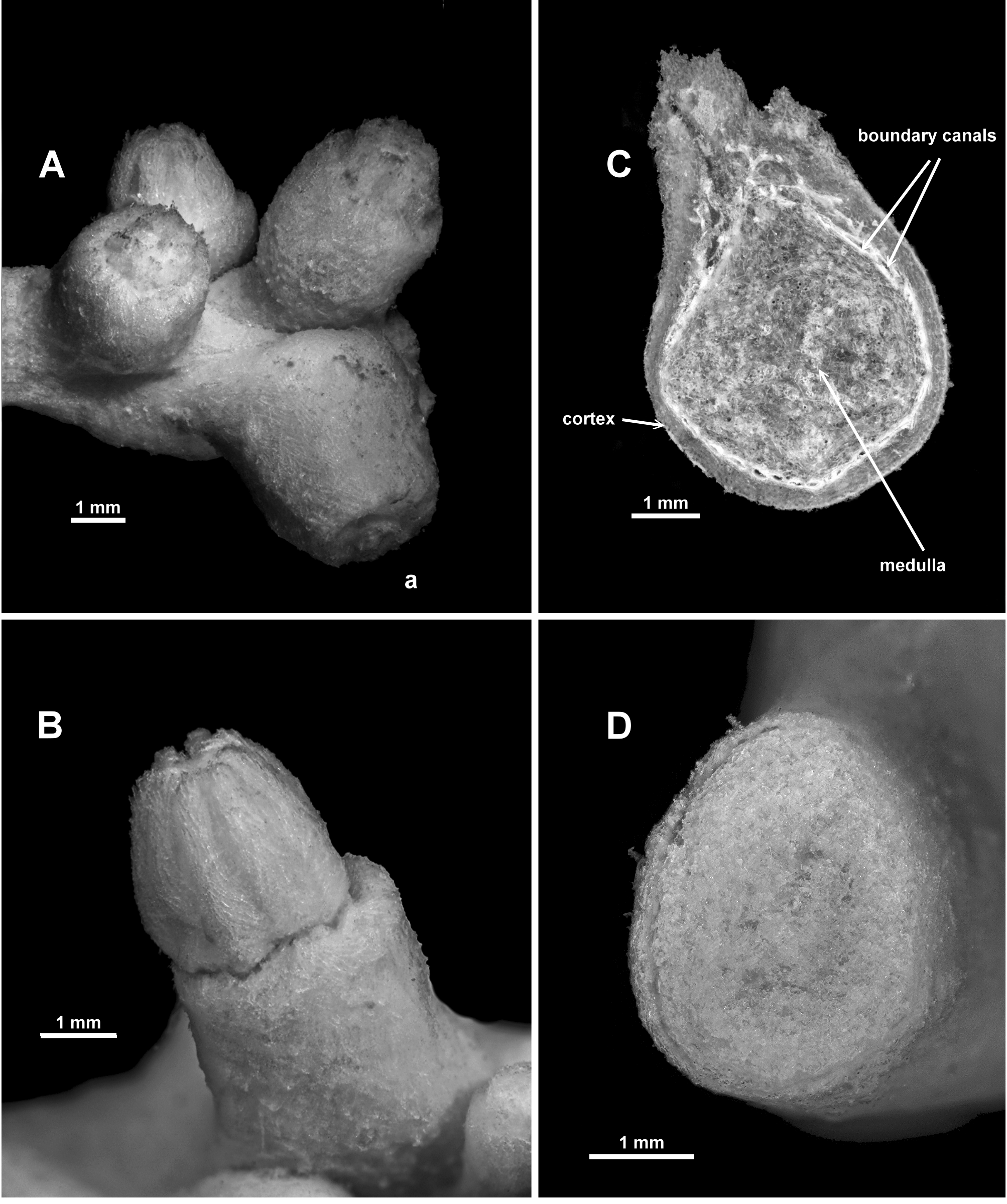
FIGURE 5. Anthothela grandiflora (Sars, 1856), holotype: A. Terminal polyp bunch (a. fully retracted polyp); B. Partly retracted polyp; C. Cross-section of decalcified medulla; D. Cross-section of medulla.

FIGURE 6. Anthothela grandiflora (Sars, 1856), holotype, sclerites: Point and collaret (a. sclerites with slightly more developed tips).
IMAGES
Gallery(27)
Occurrences with images
CITATIONS
References(22)
(2010) Rödlistade arter i Sverige 2010
Bayer, F.M. (1961). The shallow-water Octocorallia of the West Indian region. A manual for marine biologists. <em>Studies on the Fauna of Curacao and Other Caribbean Islands.</em> 12:1-373, pls. 1-23.
Breeze, H., D.S. Davis, M. Butler, and V. Kostylev. 1997. Distribution and status of deep sea corals off Nova Scotia. Ecology Action Centre. Halifax. 58 p.
Cairns, Stephen D., Dale R. Calder, Anita Brinckmann-Voss, Clovis B. Castro, Daphne G. Fautin,..., 2002: Common and Scientific Names of Aquatic Invertebrates from the United States and Canada: Cnidaria and Ctenophora, Second Edition, 2002. American Fisheries Society Special Publication 28. xi + 115.
Carlgren, O. (1940). A contribution to the knowledge of the structure and distribution of the cnidae in the Anthozoa. Kungliga Fysiografiska Sällskapets Handlingar, 51, N.F., (3): 1-62