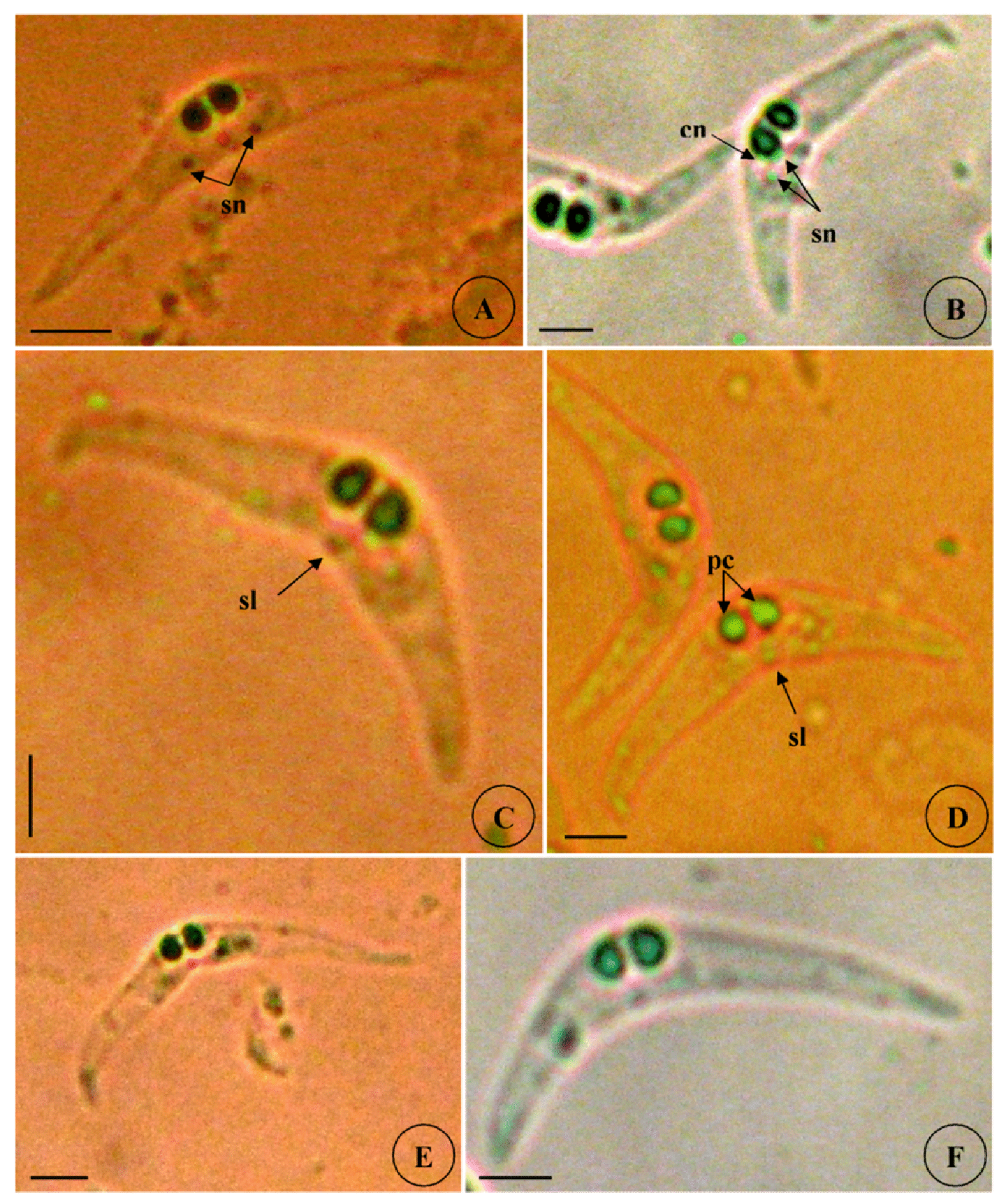
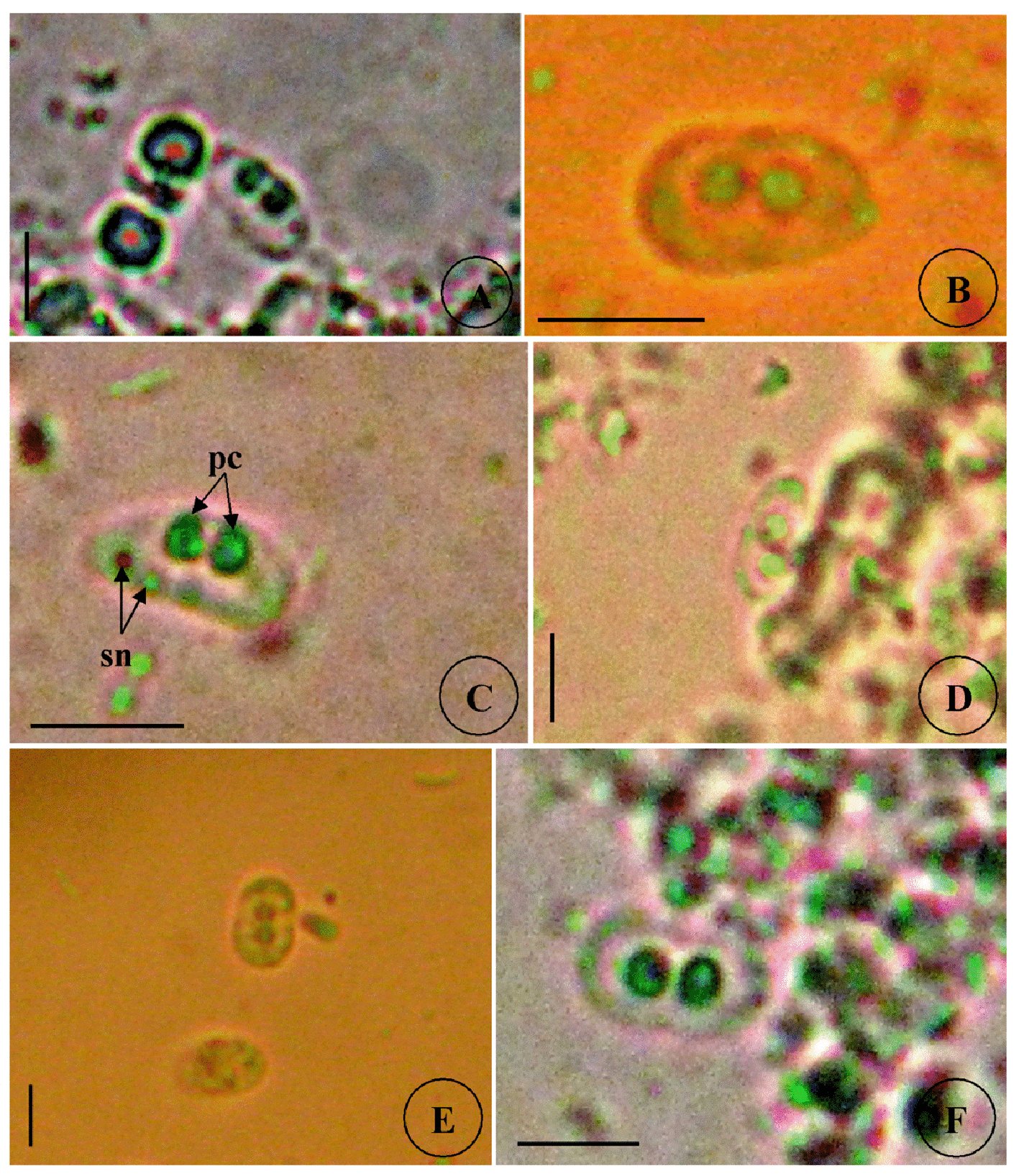
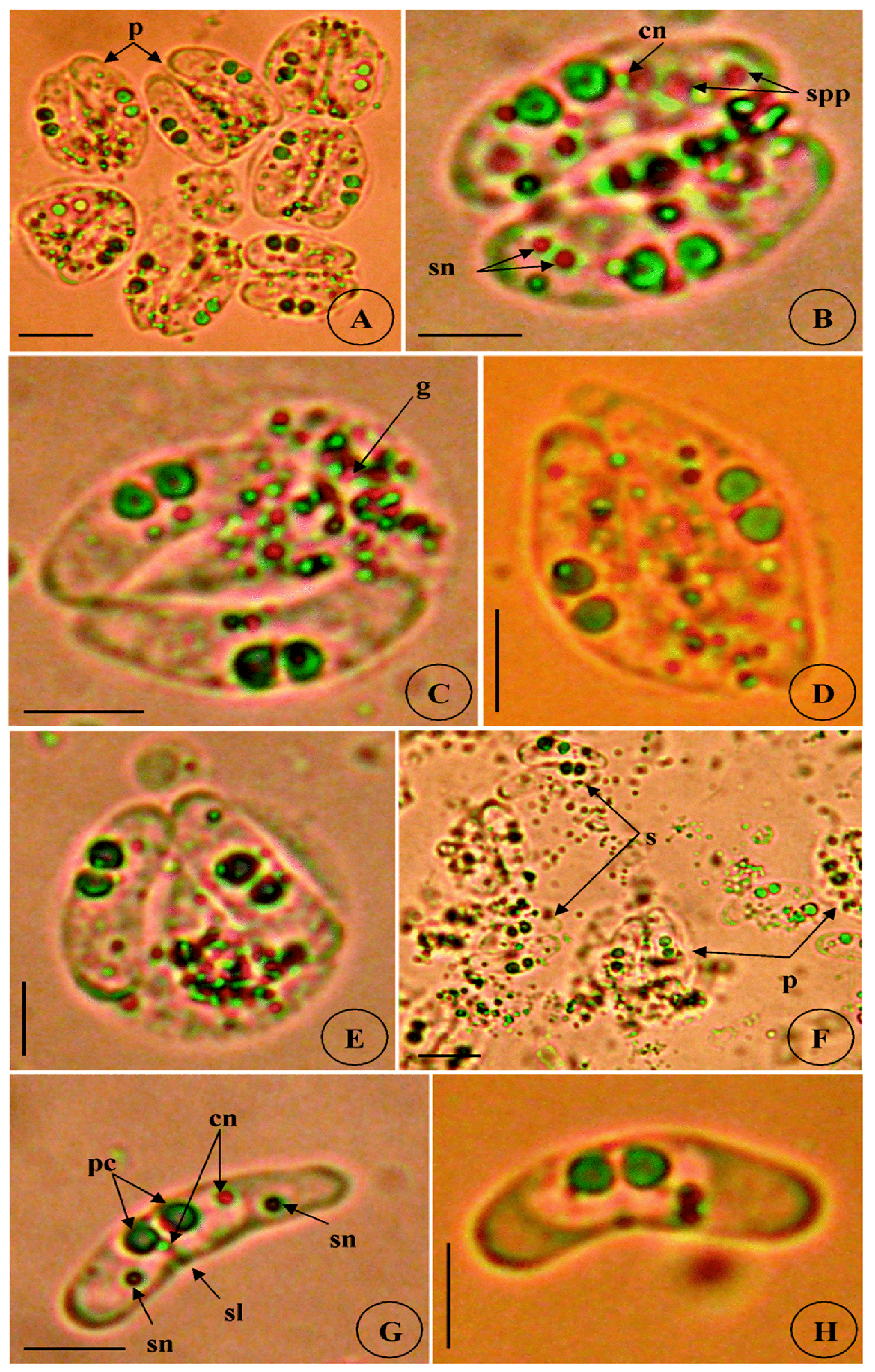
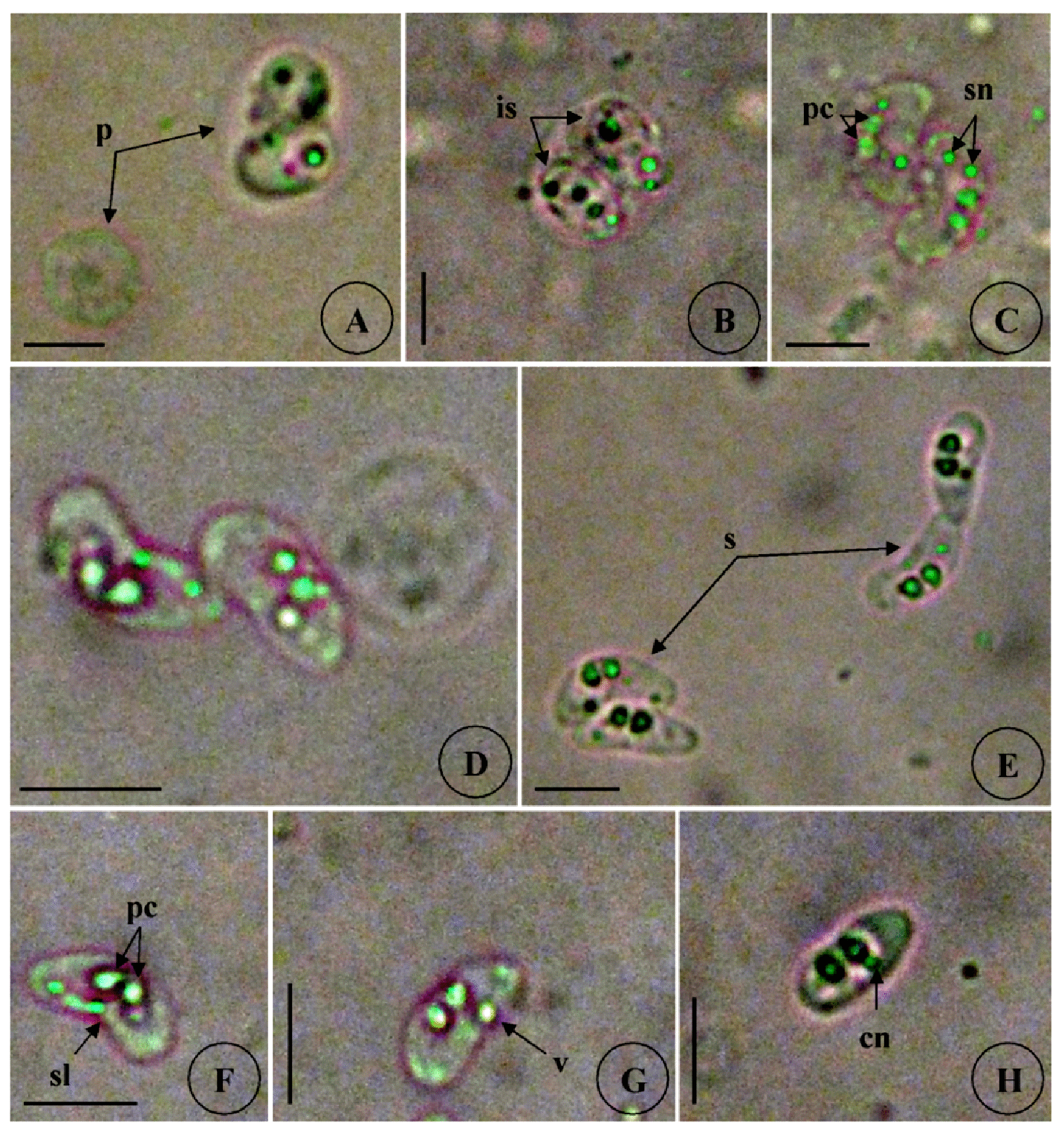
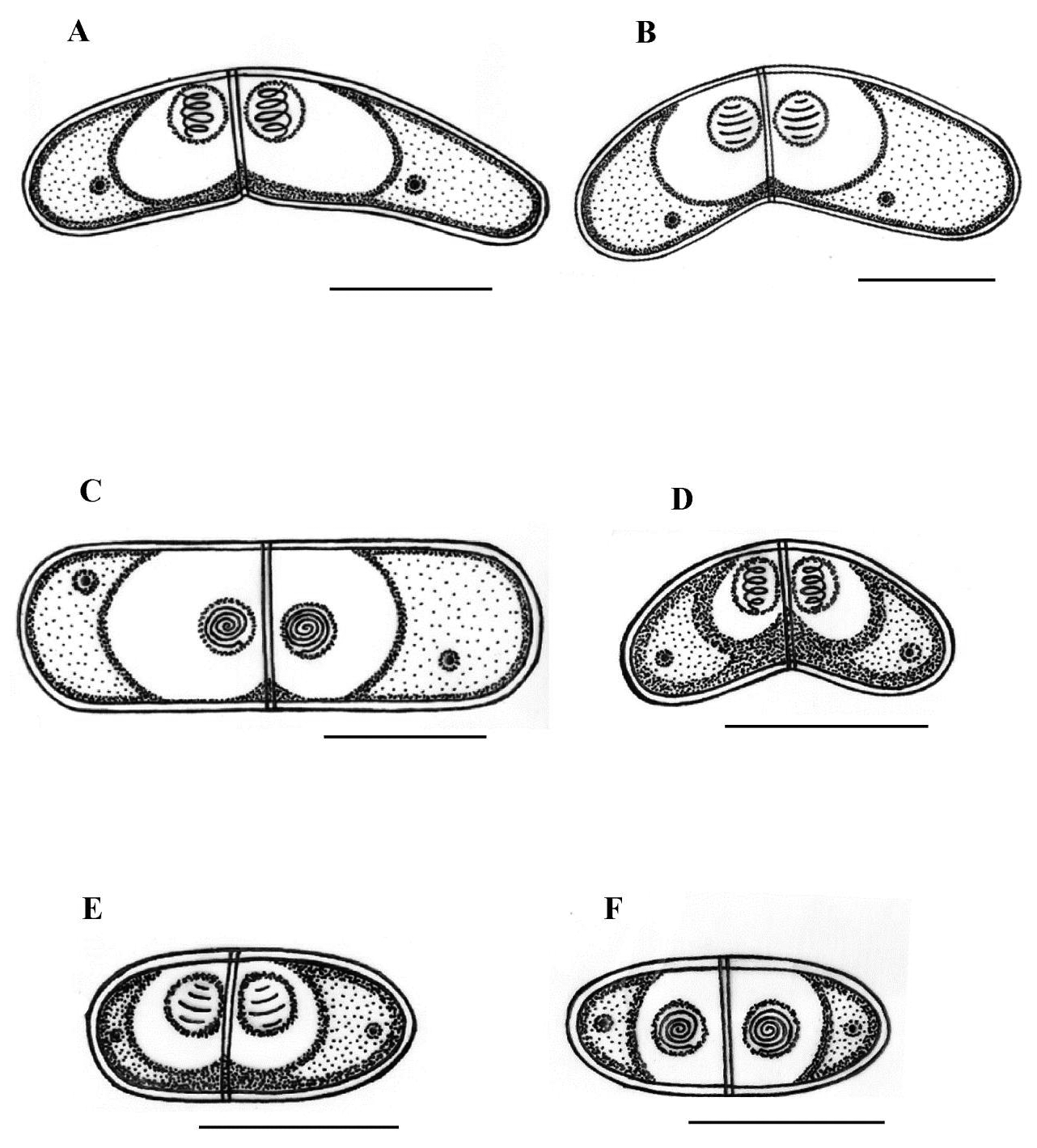
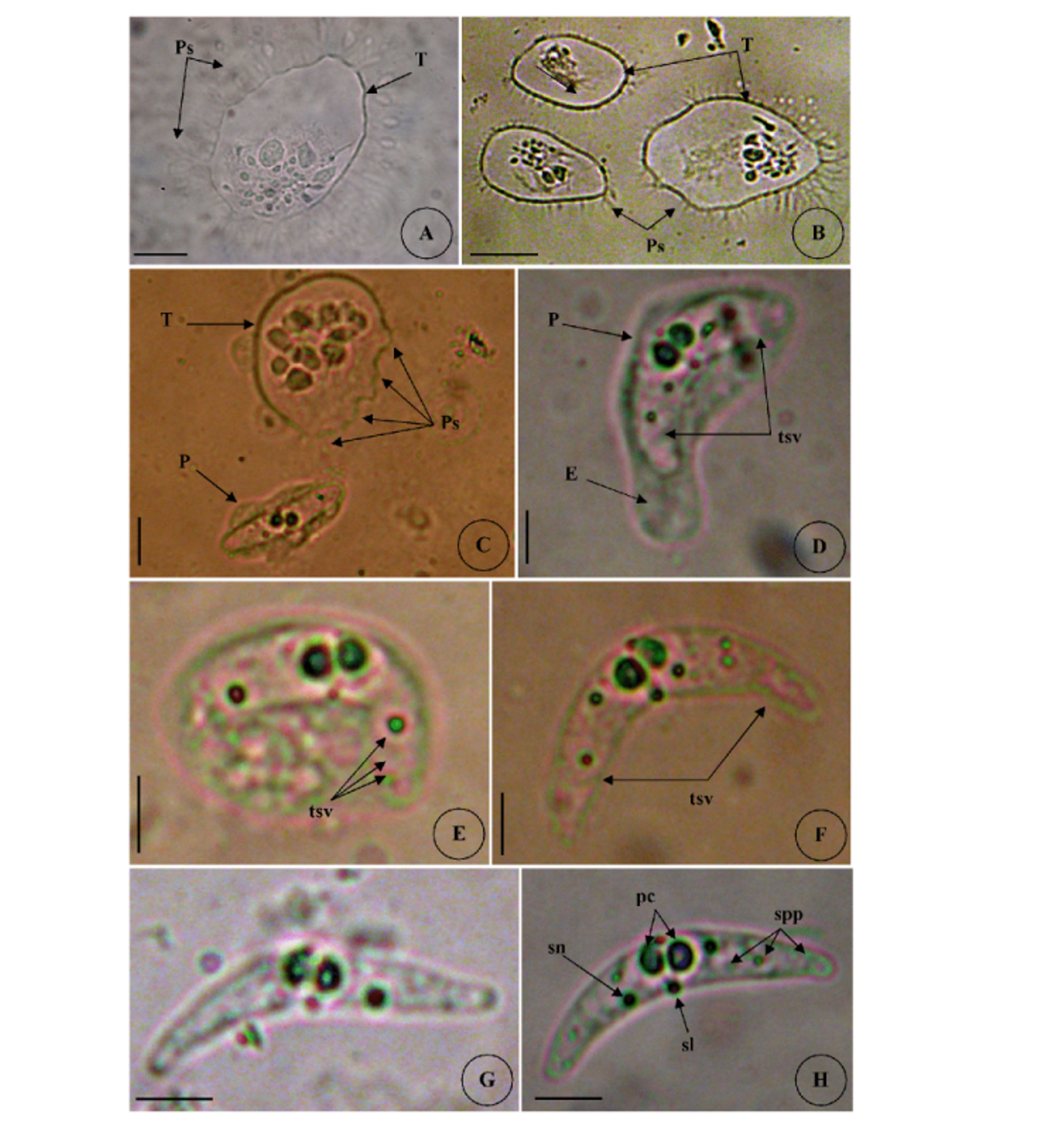
Type host: Sarpa salpa Linnaeus, 1758, goldline sea bream (Perciformes: Sparidae) Type locality: Mediterranean off Tunisia, Gulf of Tunis (36 ° 45 ’ N, 10 ° 15 ’ E). Site of infection: Within gall bladder Prevalence: The overall prevalence is 8.8 % (29 / 330) (Fig. 9). 0 % (0 / 120) fish infected in the Bay of Bizerte. The infection was confined only at Gulf of Tunis with prevalence 13.8 % (29 / 210) distributed as following, 03 / 2012: 16.7 % (5 / 30); 04 / 2012: 20 % (6 / 30); 05 / 2012: 10 % (3 / 30); 06 / 2012: 10 % (3 / 30); 07 / 2012: 20 % (6 / 30); 08 / 2012: 10 % (3 / 30); 05 / 2013: 15 % (3 / 20); 06 / 2013: 0 % (0 / 10) (see Table 4). Mean intensity: 98.5 ± 25.7 spores / infected fish (+++++) (Fig. 10) (see Table 4). Type-material: Digitized photos of syntype spores were deposited in the parasitological collection of the Museum National d’Histoire Naturelle (MNHN), Paris, Coll. No. ZS 127. Description Vegetative stages. Trophozoïtes were sub-spherical to pyriform (Figs. 5 A, B), floating freely in the bile. Some live trophozoïtes were seen with a long pseudopodia, extending on all it periphery and containing numerous refractile granules and inner generative cells (Fig. 5 A, B). Monosporic plasmodia (n = 30), measuring 23.1 ± 3.25 (18 – 27) µm in length and 38.94 ± 5.72 (32 – 48) µm in width. Each plasmodium presented one spore which the extreme part of each shell valves are twisted and surrounded with very granular endoplasm (Figs. 5 D, E). Spores (n = 30 fresh spores). Spores typical of the genus Ceratomyxa. Mature spores were elongated, arcuate to crescent-shaped in sutural view (figs. 7 A – B, 8 E) measuring 9.73 ± 0.63 (9 - 10.5) µm in length and 40.32 ± 3.83 (35 – 45) µm in thickness. Posterior angle was concave 150.2 ± 2.9 (146 – 155 °). The valves were almost equal with rounded ends and smoothly ovoid in lateral view. Anterior and posterior margins of shell valves tapered gradually to the end. A binucleate sporoplasm contained numerous sporoplasmosomes and occupied most of the spore cavity and separated by straight sutural line visible between the valves (Fig. 5 F, H). Polar capsules were pyriform 4.2 ± 0.2 (4 – 4.5) µm in length and 3.51 ± 0.39 (3 – 4) µm in width (n = 30). The polar filament formed four to five turns arranged along the longitudinal axis of the capsule. Taxonomic affinities Among the Ceratomyxa spp. reported from the Mediterranean Sea and throughout the world and under consideration of the morphological and measures characteristics of the studied species, several species seem superficially closer to the current finding. These species are: C. arcuata Thélohan, 1892 described in different hosts and localities (see Table 5). C. elongata (syn. C. lepidopusi, see Gunter & Adlard 2010), C. subtilis Meglitsch, 1960 found in the gall bladders of Lepidopus caudatus (Euphrasen, 1788) and Coelorhynchus australis (Richardson, 1838) respectively from New Zealand waters, C. rohdei Moser, Kent & Dennis, 1989 found in the gall bladder of Petroscrites fallax (Smith-Vaniz, 1976) from Australia, C. durusa Aseeva, 2003 described from the gall bladders of Limanda aspera (Pallas, 1814) and L. herzensteini (Jordan & Snyder, 1901) in Russia (Sea of Japan) and C. anko Freeman, Yokoyama & Ogawa, 2008 found in the gall bladder of the Lophius litulon (Jordan, 1902) from the Pacific coast of Japan (see Table 2). Comparative study of morphological and morphometric characteristics of the spores and polar capsules with the mentioned above species shows that the present species distinguishes from C. arcuata by having thicker spores with two shell valves narrower toward their ends compared to those of C. arcuata. The spores of C. elongata and C. durusa appear to be much thicker than those of isolate species and their polar capsules are ordinarily spherical on the contrary to the present form. For C. subtilis, only the shape of spore is closely similar to recent species, however no measurements overlap between both species. The spores of C. rohdei are shorter in length and possess two spherical polar capsules while those of the current species are pyriform. the recent finding is separated from C. anko by having two polar capsules pyriform while those of C. anko are typically spherical. Besides, the valves of the present isolate are narrower than those of C. anko. In light of these differences with closely related species, we believe that present myxozoan under study found infecting the gall bladder of S. scriba, is a morphologically a distinct species reported by the first time in S. salpa from the Mediterranean Sea. Ecological notes During this study, infection by Ceratomyxa sp. 2 was observed only at Gulf of Tunis. The overall prevalence is 8.8 %. This myxosporean has a parasitic status as satellite species. Infection by C. sp. 2 started from March to August. Prevalence and mean intensity values fluctuated with months and the maximum rate of infection was noted in April and July with 20 % (see Table 4). Overally, the mean intensity was moderate with 98.5 ± 25.7 spores per infected fish (Fig. 10). Species Host (s) Localiity Spore Polar capsule Length Thickness Length Width Ceratomyxa sp. 2 Sarpa salpa Tunisia (Gulf of 9.73 (9 – 10.5) 40.32 4.2 3.51 (Present study) Tunis) (35 – 45) (4 – 4.5) (3 – 4) Type host: Sarpa salpa Linnaeus, 1758, goldline sea bream (Perciformes: Sparidae) Type locality: Mediterranean off Tunisia, Bay of Bizerte, Tunisia (37 ° 20 ’ N, 9 ° 53 ’ E). Site of infection: Within gall bladder Prevalence: The overall prevalence is 3 % (10 / 330) (Fig. 9). None fish caught from Gulf of Tunis was parasitized with C. sp. 3. The infection was recorded only at Bay of Bizerte with prevalence 8.33 % (10 / 120) distributed as following, 03 / 2013: 0 % (0 / 30); 04 / 2013: 0 % (0 / 30); 05 / 2013: 33.3 % (10 / 30); 06 / 2013: 0 % (0 / 30) (see Table 4). Mean intensity: 35.5 ± 8.4 spores / infected fish (++++) (Fig. 10) (see Table 4). Type-material: Digitized photos of syntype spores were deposited in the parasitological collection of the Museum National d’Histoire Naturelle (MNHN), Paris, Coll. No. ZS 128. Description Vegetative stages. Trophozoïtes (n = 30) were pyriform, floating freely in the bile and measured 19.6 ± 2.05 (16.9 – 22.8) µm in length and 32.68 ± 2.57 (29.5 – 36.4) µm in width. Disporous, each plasmodium contained two identical spores with fine granules (Fig. 6 A, B). Spores (n = 30 fresh spores). Spores typical of the genus Ceratomyxa. Mature spores were narrowly crescentshaped and transversely elongate, with anterior margin slightly convex and posterior slightly concave in sutural view (Figs. 6 C – E, 8 F), measuring 7.4 ± 0.8 (6.5 – 8.5) µm in length and 30 ± 1.8 (28 – 33) µm in thickness. Posterior angle was slightly concave to straight 168.5 ± 4.2 (162 – 172 °). Two shell valves equal in size and straight sutural line visible between valves (Figs. 6 C – F). The most attractive feature was a binucleate sporoplasm not filling all the spore cavity so that the space around each polar capsule was completely free (Figs. 6 C – F). Polar capsules were typically spherical, 3 ± 0.41 (2.5 – 3.5) µm in length and 3 ± 0.41 (2.5 - 3.5) µm in width (n = 30). The polar filament coiled with four to five turns, situated perpendicularly to the longitudinal axis of the capsule. Taxonomic affinities In Mediterranean Sea, Ceratomyxa sp. 3 shows morphological resemblances to four species of Ceratomyxa which are C. pallida Thélohan, 1895 described from the gall bladders of S. salpa and B. boops in France, C. puntazzi and C. sp 1 Alama-Bermejo, Raga & Holzer, 2011 described from the gall bladders of Diplodus puntazzo (Walbaum, 1792) and Sparus aurata (Linnaeus, 1758) respectively in Spain and C. filamentosi Kalatzis, Kokkari & Katharios, 2013 described recently from the gall bladder of Aulopus filamentosus (Bloch, 1792) in Greece (Cretan sea) (see Tables 3 and 5). The comparison between mentioned above species and current isolate proves that all the measurements of C. pallida, C. puntazzi, C. filamentosi and recent species overlap, however, the difference between all of these species and the present finding is transflated in the shape and the place of sporoplasm within current spores whose does not occupy the entire spore cavity. Besides, the only species that has a disporic plasmodia as recent form, is C. puntazzi that characterized by plasmodia with very granular protoplasm and motile pseudopodia-like filopodia which are both lacking at current species. In addition, the present finding differs from C. sp. 1 ex S. aurata by having spores larger in length and much thicker. In others areas around the world, the recent species is superficially similar to C. moenei Meglitsch, 1960 found in the gall bladder of Polyprionum moene (Phillips) from New Zealand, C. seriolae Yokoyama & Fukuda, 2001 found in the gallbladder of Seriola quinqueradiata (Temminck & Schlegel, 1845) from Japan, C. azonusi Asseva, 2003 found in the gall bladder of Pleurogrammus azonus (Jordan & Metz, 1913) from Russia (Sea of Japan) and C. milleri Gunter, Whipps & Adlard, 2009 found in the gall bladder of Lutjanus fulviflamma (Forsskål, 1775) from Great Barrier Reef (Australia) (see Table 3). From the first evaluation, it was clearly that recent isolate is differentiated from all 4 species by the shape and place of sporoplasm inside the spore. Furthermore, C. moenei has a posterior margin that is more strongly concave than that of the present species. C. azonusi differs from current finding by having a pyriform polar capsules while those of the present species are typically spherical. The spores of C. milleri appear to be thinner than those of the current species besides no measurements overlap between both species. The spores of C. seriolae are shorter in length and have a smaller polar capsules compared to those of recent form. Therefore, taking account all the revealed differences with closely related congeneric species, the present myxosporean has a dissimilar morphological characteristics and it designated as a different species, not previously identified in S. salpa from the Mediterranean Sea. Ecological notes During this study, the overall prevalence is 3 %. This myxosporean has a parasitic status as scarce species. Infection by Ceratomyxa sp. 3 was restricted only in Bay of Bizerte and this species was detected only in May with prevalence 33.3 % and mean intensity 35.5 ± 8.4 spores per infected fish (see Table 4). Species Host (s) Locality Spore Polar capsule Length thickness Length Width
New observations on Myxozoa of the goldline sea bream Sarpa salpa L. 1758 (Teleostei: Sparidae) from the Mediterranean coast of Tunisia