Molgulidae
kulsjöpungar(+8)
GBIF:9304
0
Descendants
0
Synonyms
0
Children
0
Species
0
Genera
PROFILE
Species Profile
Habitat
ABOUT
Descriptions(1)
Molgulidae
Molgula euplicata Herdman, 1923 (Figures 31, 32)
Herdman, 1923: 15. Monniot & Monniot 1983: 102 and synonymy; 1994: 34.
Stations (events when several trawling operations per station): 27(33)-49A-57-59.
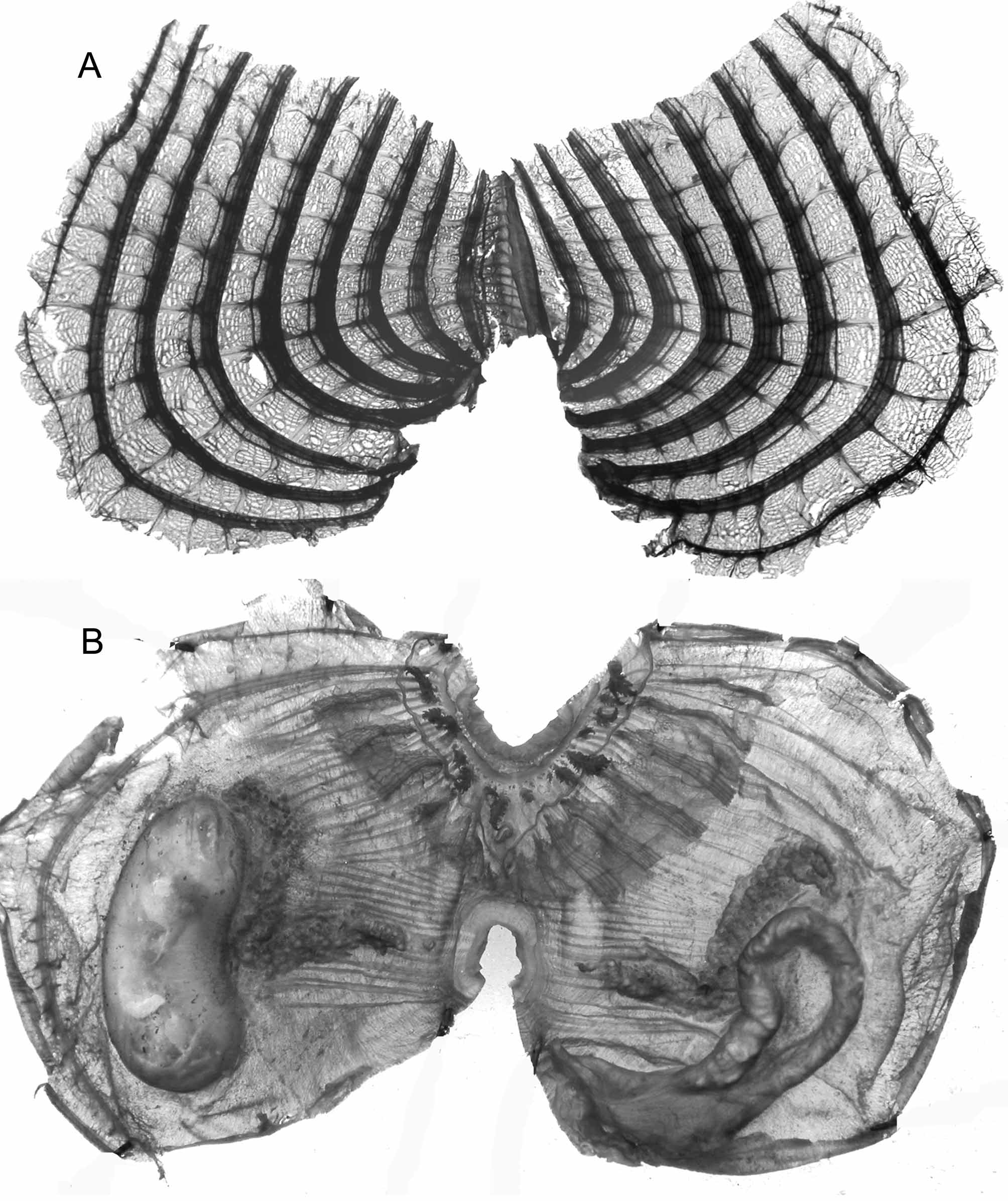
Numerous specimens were collected between 450 and 1250 m depth. They seem to be free living on the sediment. They are globular, covered with sand and hair-like processes. The sessile siphons, difficult to detect, are distant of half the body length. The soft tunic is easily removed from the body wall. The edge of the oral siphon has 6 pointed lobes. In addition to the sphincters, strong muscular ribbons start from the siphons but do not reach the ventral body side (Figs 31 B, 32A), and a felting of criss-crossed thin fibres are distributed on the whole body wall. Twelve to 14 flat and short oral tentacles, with only primary branches alternate with 2 orders of smaller ones, the smallest being button-like. The dorsal tubercle is clearly protruding, opening in a C and located into a wide curve of the peripharyngeal band (Fig. 31 B, 32A). The dorsal lamina is short (Fig. 31 A) with a dentate edge in its posterior part. The branchial sac has always 9 folds on the right side but 8 or 9 on the left side, the 9th fold sometimes reduced to a longitudinal vessel (Fig. 31 A). There are 12 to 14 longitudinal vessels on the larger folds. There are no exo-infundibula (Fig. 32 B).The gut is curved in a simple loop, and is included into the body wall (Fig. 31 B, 32A). The anus opens close to the oesophagus with a plain or slightly dentate edge. The hepatic gland does not protrude in papillae but remains indistinct on the stomach. The kidney is voluminous and arc-shaped (Fig. 31 B, 32A), it contains a solid inclusion with a laminated structure. On each side a long sinuous gonad is entirely linked to the body wall; the central ovary is fringed with testis vesicles. Multiple male papillae protrude on the internal side of the ovary (Fig. 32 A, C).
This species is well defined by its 9 branchial folds, and the long sinuous gonads with many male papillae.
Sequences from specimen S3 MOL.A 387a (BOLD: ASCAN046-10) and specimen S3 MOL.A 386a (BOLD: ASCAN045-10) show 1.06% divergence. No close hit in BOLD (best: 70.76%).
In spite of its wide Antarctic distribution, previous descriptions were not detailed. In Terre Adélie the specimens were found from 400m but particularly abundant from 1000 to 1250 m depth.
Molgula pedunculata Herdman, 1881 (Figures 33, 34, 35)
Herdman, 1881: 234. Monniot & Monniot 1983: 98 and synonymy. Primo & Vazquez 2007: 1806. Tatian et al. 1998: 211.
Stations (events when several trawling operations per station): 1-3-5-11 (424)-11(429)- 12-14-16 A-17-20-26A- 30(66)-47-49A-51-70-71-79-86E.
The large bodies are erect on a peduncle of variable length. In a sparse distribution the peduncle is short (Fig.33 A) and the internal organs thicker (Fig. 34 A) when in crowded populations the peduncle is long (fig. 35) and the organs more spread out on the body wall (Fig. 33 C). Small individuals have short papillae on the whole tunic (Fig. 33 B), in largest specimens papillae only occur on the siphons. In Terre Adélie the body reaches 17 cm in length, the longest peduncles are often broken in trawls. The musculature (Fig. 33 C, 34A) comprises: the siphonal sphincters, thin circular fibres included into the large velum in each siphon, ribbons issued from the siphons spreading on each side until the middle of the body, and thinner transverse fibres. The body wall extends into the peduncle with muscles. The branchial sac has 7 folds. The number of longitudinal vessels increases with the body size and the folds become more spaced (Fig. 34 B). The dorsal lamina is high and joined to the branchial folds on the left side. The gut loop has a different shape in specimens with a long or a short peduncle, with intermediate situations. In specimens with a long stem the gut is in vertical position in a long straight loop (Fig. 33 C), when in individuals with short peduncle the gut loop is curved and occupies a larger part of the left side (Fig. 34 A). In both cases the rectum is fused to the stomach on a long distance, leaving a narrow lumen. The hepatic gland is voluminous and covers the whole stomach. The end of the rectum and the smooth edged anus are strongly attached to the dorsal lamina. There is one gonad on each side (Figs 33 C, 33A), oval and more bulky in stocky individuals and elongate in thinner specimens. In any case the testis vesicles are in small groups along the central ovary. A series of short male papillae open along the internal side of the ovary. The oviduct is short. The proximal part of the left gonad is always fused with the intestine; the proximal tip of the right gonad is joined to the kidney. The arc-shaped kidney (Figs 33 C, 34A) contains dark spherical granules.
Sequences from specimen S3 MOL.A 378a (BOLD: ASCAN048-10) and specimen S3 MOL.A 373a (BOLD: ASCAN047-10) display 0.38% divergence. Both sequenced specimens have rather long peduncles. No close hit in BOLD (best: 75%).
Molgula pedunculata is widely distributed in the Antarctic and Sub-Antarctic waters from shallow depths to 1000 m, and can constitute dense spectacular populations as shown by pictures in situ (Fig. 35)
Molgula riddlei Monnoit n. sp. (Figures 36, 37)
The species is named after Dr. Martin Riddle, Chief Scientist on board the RSV Aurora Australis.
Station: 80.
The single specimen 15 mm in diameter is round, covered with sand and short hairs. The siphons could not be seen externally. After removing the tunic the siphons are sessile, well apart with a thick wall, the body wall is thinner. The musculature is divided in several sectors: strong sphincters around the siphons, 2 separate fields of ribbons issued from each side of both siphons, and very short transversal fibres arranged in 2 parallel lines on each side (Fig. 36). Thinner transverse fibres, which do not extend over the body sides, are dorsal between the siphons. Slen- der oral tentacles are moderately bushy with primary and secondary branches. The peribranchial groove has 2 crests. The dorsal tubercle opens in a vertical slit. The dorsal lamina is high, short, smooth edged (Fig. 37). The 7 branchial folds do not recover each other. There are no longitudinal vessels between the folds (Fig. 37). The branchial formula is:
LE - 8- 11 – 11 – 10 – 11 – 10- 10 –DL – 10 -10 – 11 – 11 - 11- 11- 8 –E R
There are no intermediate stigmatic spirals but only regular infundibula. The branchial folds join the retropharyngeal band. The gut occupies a small part of the left side (Fig. 37). The round stomach is covered with round low hepatic papillae. The narrow intestine draws a large loop and a deep secondary bend, linked to the stomach and oesophagus, the long rectum attached to the body wall ends in a bilobed anus. A thin pyloric canal crosses the primary intestinal loop. The kidney is voluminous egg-shaped (Fig. 36). There is one gonad on each side (Fig. 36). The left one is oval located in the secondary intestinal loop; the right one is curved pressed against the kidney. The wide oviduct is apical full of eggs (220µm in diameter) and directed toward the atrial opening. The male vesicles form an irregular mass around the ovary. The sperm duct ends in a long tubular papilla (Fig. 36).
36. Molgula riddlei n. sp. body ventrally opened, scale bar = 5mm.
Among Antarctic and Sub-Antarctic sandy Molgula species with 7 folds on each side Molgula riddlei can be compared to Molgula enodis (Sluiter, 1912) but the latter has less branchial vessels and a different musculature. Molgula mortenseni (Michaelsen, 1922) from New Zealand has internal papillae on the branchial vessels and a short rectum.
Molecular results and discussion
While amplifying and sequencing COI is generally straightforward, problems arise for at least some parts of many taxonomic groups, and can bias the representation of taxa in the sequence databases. We found that relatively few ascidian species are represented in BOLD, probably because of technical difficulties in producing DNA sequences for COI. The depth of sequence divergence makes the use of several pairs of primers necessary. Even so, the COI barcoding of Ascidiacea is technically challenging, and sequences could not be obtained for all specimens and all species. The problem is COI-specific, as other markers could be amplified without problem from the same DNA extracts (data not shown). Moreover, in the two third of our samples (including distantly-related species), replication slippage during PCR on a stretch of ten Ts (around position 150 of the Folmer region) causes a degradation in sequence quality after the poly-T. At least two PCRs had to be sequenced in both directions before obtaining reliable sequences. These technical difficulties might cause considerable troubles for the use of COI barcoding on Ascidiacea.
Thirty seven sequences were obtained for 26 species (see descriptions part for each species), out of 49 specimens and 31 species tested. The sequences could be obtained for the type specimens of two of the three herein described new species and for the paratype of the third. The sequences contained no stop codon when analysed using the Ascidian mitochondrial code as implemented in CodonCode Aligner.
In our Ascidiacea sampling as well as in the sequences available in GenBank, the base composition is heavily skewed towards A and T bases: in our dataset, the average over all sequences calculated with MEGA (Tamura et al. 2007) is 40.7% T (min: 34.1%; max: 45.4%), 15.5% C (min: 11.8%; max: 21.1%), 27% A (min: 20.3%; max: 33.1%), 18.8% G (min: 13.8%; max: 24.2%). This might explain the very low primer hybridization temperature (40°C) necessary for successful PCR on the COI gene whatever the primer pair used.
Intraspecific divergences ranged from 0% to 20.77%). These values might be underestimations, as we had at most sequences for two specimens per species. Most intraspecific divergences were below or around 1%, but the sequences for the two specimens of Aplidium balleniae diverged by 2.46%, and this divergence went to 11.79% for Pyura bouvetensis and to 20.77% for Didemnum biglans . Interspecific distances were generally above 6.9% (mean 22.39%), except for the two Distaplia specimens ( D. colligans and D. cylindrica), which differed only by 0.17%. The anatomy of these two species is very similar, and only the shape of the colonies differs slightly. When analyzing the other 751 Ascidiacea sequences available in BOLD, similar high intraspecific values and low interspecific values are found. The level of maximal and minimal inter and intraspecific variability for COI can vary even between closely related Antarctic species (Dettai et al. 2010). Further studies combining multiple sequence datasets and morphology, and including a larger number of specimens are needed to explore both interspecific and intraspecific variability for Antarctic ascidians.
No sequences from outside our dataset provided close hits in the complete BOLD database. A search using the species names showed that for most species from our dataset all sequences come from our study (24 species out of 26), which therefore provides a much needed reference dataset for Antarctic ascidians.
This tunicate collection is unique in several respects. Three new species were described. Among these, Molgula riddlei was found associated to dense coral-sponge communities at the head of the shelf cutting Cuvier Canyon (Post et al. 2010b) a location which was designated as a VME (Vulnerable Marine Ecosystem) by the CCAMLR. During the CEAMARC cruise, video and still footage revealed for the first time that Molgula pedunculata form spectacular monotypic communities associated to iceberg scours (Fig. 35). This species is likely to be part of the recovery process of faunal assemblages after iceberg disturbance on the East Antarctic shelf. This is also the first time that so many specimens suitable for DNA sequencing were collected from the East-Antarctic shelf. The combined molecular and morphological approach provided unexpected insight in Ascidiacea taxonomy. This alone will have strong repercussions on our understanding of many crucial aspects of Antarctic scientific and environmental issues, such as circumpolarity, shelf-slope faunal interactions, eurybathy, and patchiness of benthos distribution.
Many species included in this study are thought to be circumpolar in distribution, with sometimes extension of their geographical range to the Sub-Antarctic regions. However, molecular results show that some taxa may be more diverse than previously thought (e.g. Pyura bouvetensis and Didemnum biglans). The tunicate biodiversity is therefore likely to be higher and some circumpolar and eurybathic taxa will certainly prove to be of more restricted distribution ranges. Two species, Bathypera hastaefera and Culeolus antarcticus, known from abyssal depths, were found at much shallower depths on the shelf of the study area. These results indicate that some tunicates may have found shelter in slope refuges during the last glacial maximum. This is also indicated by the high rate of eurybathy among the studied species. Species composition of adjacent stations [i.e. 37(146) & 37(147), 36(68) & 36(297), 32A & 33A, 65(318) & 65(322), 27(33) & 27(45) & 27(46), 6(99) & 6(103), 11(424) & 11(429)] differ notably. This shows that the sorting/sampling effort was probably not as homogenous as was done for fish, crustacean and crinoids. However, it may also indicate that the different habitats sampled are excessively patchy and that pseudoreplicate stations are likely to display different faunal associations.
This unique Ascidiacea collection emphasizes the need to further investigate the systematic of Antarctic tunicates. It also stresses on the fact that some species may reveal to be model or key organisms to study the dynamic of faunal communities on the shelf and slope around the continent.
Export occurrence data
Darwin Core Archive (ZIP)
GEOGRAPHY
Distribution Map
REGIONS
Geographic Distribution(4)
DATA
Occurrence Datasets
Common names used for this species across different languages and regions. Available in 5 languages and 1 country. 1 preferred.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide. This taxon has names in 5 languages. 1 name preferred.
CLASSIFICATION
Taxonomic Classification Tree

NOMENCLATURE
Synonyms(1)
RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(6)
FIGURE 31. Molgula euplicata. A, branchial sac; B; body ventrally opened.

FIGURE 32. Molgula euplicata: A, body ventrally opened, scale bar = 10 mm; B, part of the branchial sac; C, detail of the left gonad.
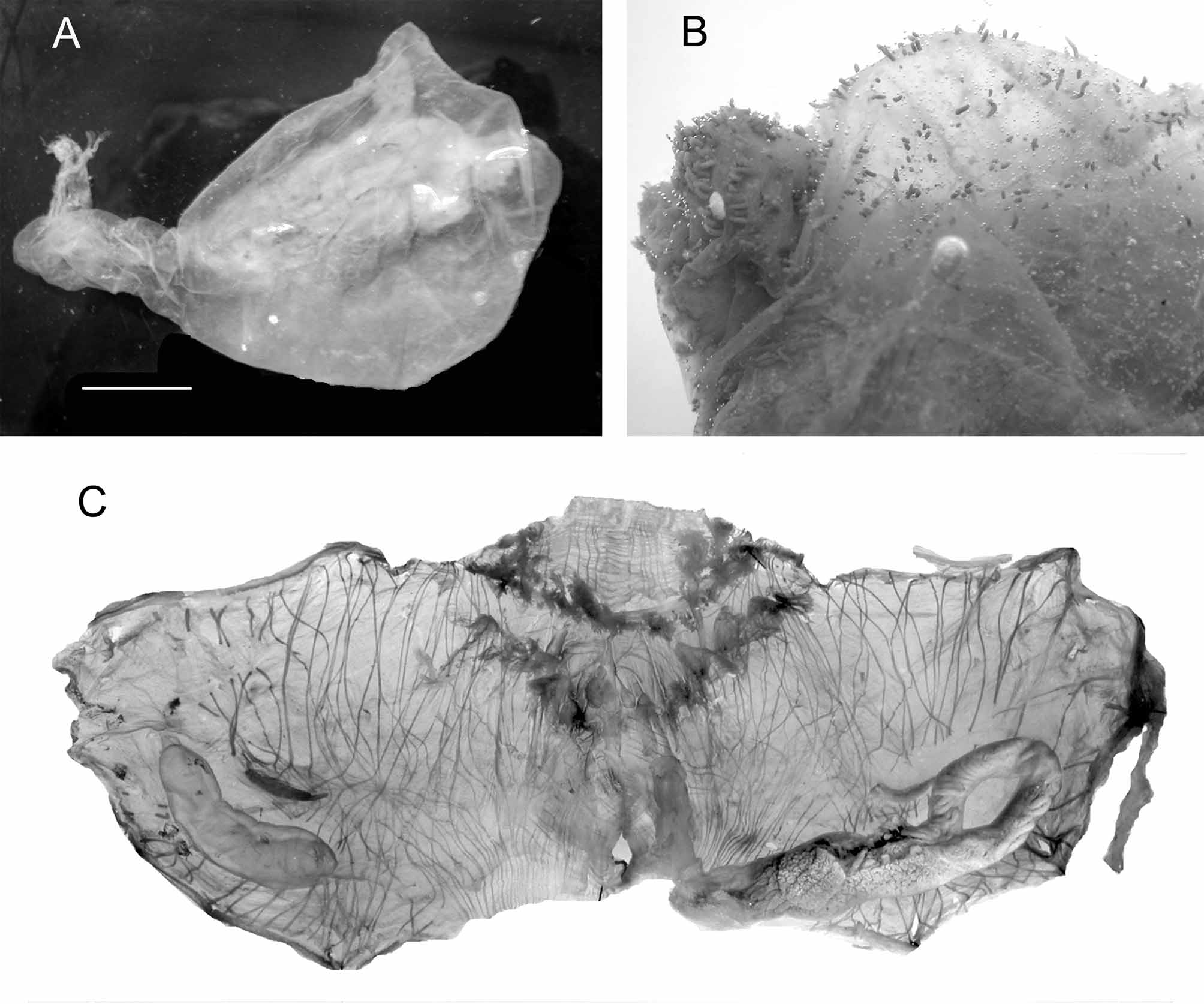
FIGURE 33. Molgula pedunculata: A, specimen with short peduncle; B, papillae on the tunic; C, specimen with long peduncle ventrally opened.
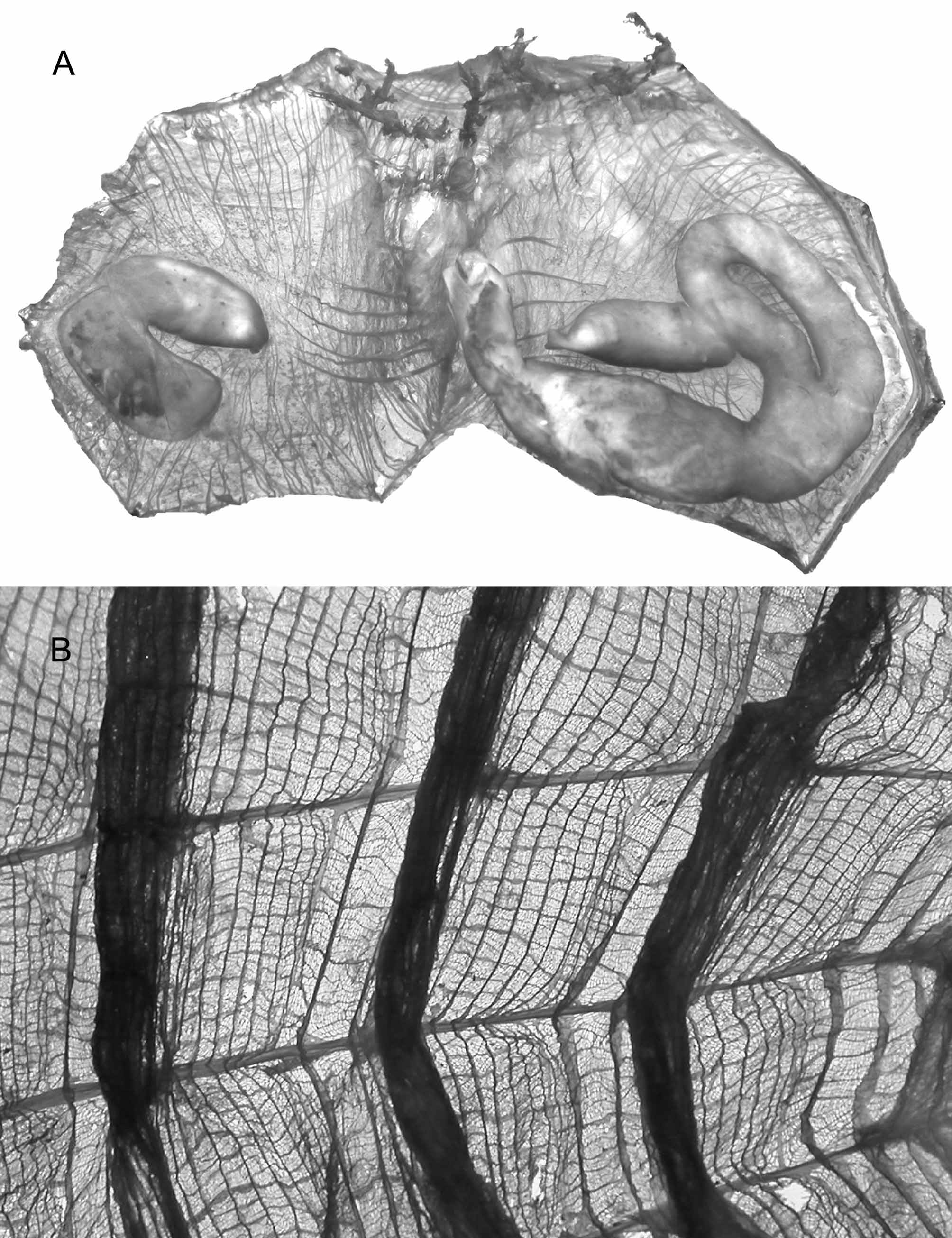
FIGURE 34. Molgula pedunculata: A, body of a specimen with short peduncle ventrally opened; B, part of the branchial sac.


IMAGES
Gallery(6)
Occurrences with images
CITATIONS
References(18)
Buizer, D.A.G. (1983)
Gärdenfors, Hall, Hallingbäck, Hansson & Hedström (2003) Djur, svampar och växter i Sverige 2003. Förteckning över antal arter per familj.
Hayward, P.J. & J.S. Ryland (Eds.). (1990). The marine fauna of the British Isles and North-West Europe: 1. Introduction and protozoans to arthropods. <em>Clarendon Press: Oxford, UK.</em> 627 pp.
Hayward, P.J. & J.S. Ryland (Eds.). (1990). The marine fauna of the British Isles and North-West Europe: 1. Introduction and protozoans to arthropods. <em>Clarendon Press: Oxford, UK.</em> 627 pp.
Huber, J. L.; Da Silva, K. B.; Bates, W. R.; Swalla, B. J. (2000). The evolution of anural larvae in molgulid ascidians. <em>Seminars in Cell & Developmental Biology.</em> 11(6): 419-426.