Caudata
salamandere(+23)
GBIF:953
0
Descendants
0
Synonyms
0
Children
0
Species
0
Genera
0
Families
PROFILE
Species Profile
Characteristics
ABOUT
Descriptions(8)
ORDER CAUDATA Fischer 1813, 58
(1 family, 5 genera, 7 subgenera, 38 species, 24 endemic species). Dubois & Raffaëlli (2012) argued for using Urodela for this order. Fouquette & Dubois (2014) continued the arguments of Dubois & Raffaëlli (2012). A.M.C. Duméril (1805, 91) had used the French vernacular Urodèles for this order. Fischer used Caudati as apparently an equivalent of Urodeli of Duméril. As the CODE does not regulate order names it comes down to a matter of choice, thus I continue to use Caudata for the salamander order, in part, because Fischer used Caudati before he used Urodeli.
Export occurrence data
Darwin Core Archive (ZIP)
GEOGRAPHY
Distribution Map
REGIONS
Geographic Distribution(4)
DATA
Occurrence Datasets
Common names used for this species across different languages and regions. Available in 7 languages and 5 countries. 2 preferred.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide. This taxon has names in 7 languages. 2 names preferred.

NOMENCLATURE
Synonyms(1)
RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(2)
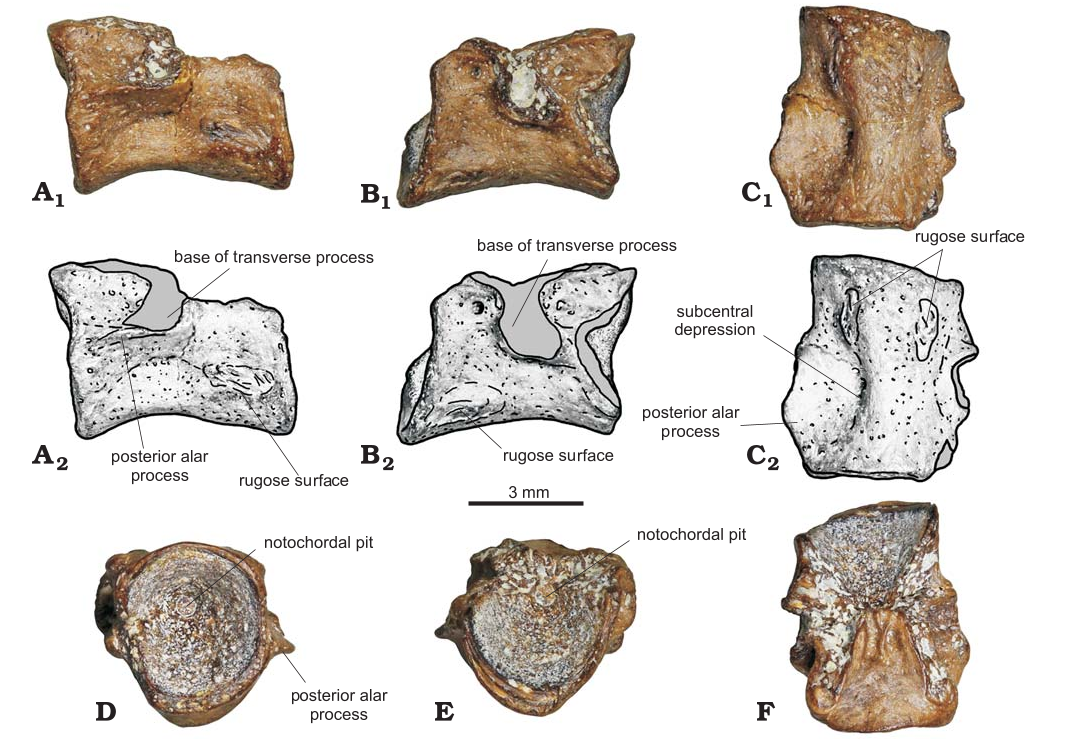
Fig.1. Fragmentary trunk vertebral centrum (LMCCE 1/4) of stem salamander Caudata, gen.et sp. indet. from the Shestakovo locality, Lower Cretaceous Aptian–Albian), Ilek Formation, Western Siberia, Russia; in right lateral (A), left lateral (B), ventral (C, anterior towards top), posterior (D), anterior (E), and dorsal (F, anterior towards top) views. Photographs (A1–C1, D–F) and interpretive drawings (A2–C2).
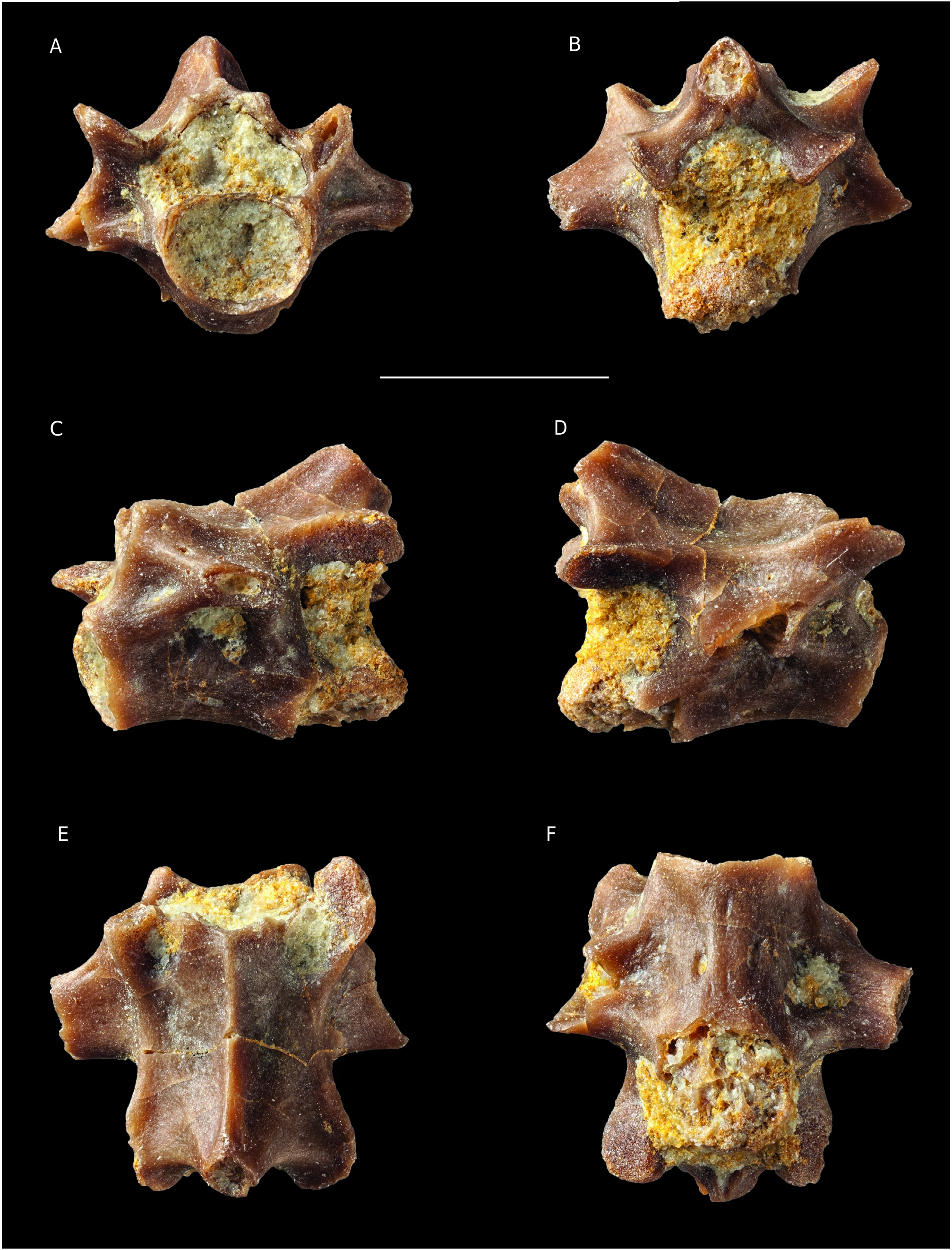
FIG. 11. — Vertebra of Caudata indet. from Angeac-Charente: A-F, (ANG M-71) in anterior (A), posterior (B), left lateral (C), right lateral (D), dorsal (E) and ventral (F) views. Scale bar: 2 mm.
IMAGES
Gallery(2)
Occurrences with images
CITATIONS
References(8)
American Museum of Natural History (2026) Amphibian Species of the World 6.2, an Online Reference
Banks, R. C., R. W. McDiarmid, and A. L. Gardner, 1987: Checklist of Vertebrates of the United States, the U.S. Territories, and Canada. Resource Publication, no. 166. 79.
Flores-Villela, Oscar / McCoy, C. J., ed., 1993: Herpetofauna Mexicana: Lista anotada de las especies de anfibios y reptiles de México, cambios taxonómicos recientes, y nuevas especies. Carnegie Museum of Natural History Special Publication, no. 17. iv + 73.
Parker, S.P. (ed). (1982). Synopsis and Classification of Living Organisms. McGraw-Hill, New York. 2 volumes.
Ruggiero, M. A., D. P. Gordon, T. M. Orrell, N. Bailly, T. Bourgoin, R. C. Brusca, et al., 2015: Correction: A Higher Level Classification of All Living Organisms. PLoS ONE vol. 10, no. 6, e0130114.