
Accipitridae
Aasvoëls en Arende(+99)
GBIF:2877
0
Descendants
0
Synonyms
0
Children
0
Species
0
Genera
PROFILE
Species Profile
Characteristics
ABOUT
Descriptions(24)
Family Accipitridae
Leptodon forbesi (IUCN: CR, Biodiversitas: Not included) Physaloptera losseni
ACCIPITRIDAE
Buteo sp.
COLEOPTERA: Trogidae
Omorgus monachus (Herbst, 1790)
UNITED STATES: Maryland: Jackson's Island, June, one specimen with the notation “ex nest buzzard” (Vaurie 1955).
Export occurrence data
Darwin Core Archive (ZIP)
GEOGRAPHY
Distribution Map
REGIONS
Geographic Distribution(4)
DATA
Occurrence Datasets
Common names used for this species across different languages and regions. Available in 47 languages and 5 countries.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide. This taxon has names in 47 languages.
CLASSIFICATION
Taxonomic Classification Tree

RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(8)
Figure 11. Accipitrid distal right humerus SAMA P.58917 in cranial (A), caudal (B), dorsal (C) and ventral (D) view. Abbreviations: CD, condylus dorsalis; CDCS, condylus dorsalis caudal scars; CV, condylus ventralis; EV, epicondylus ventralis; FB, fossa brachialis; FO, fossa olecrani; II, incisura intercondylaris; MECR, m. extensor carpi radialis insertion scars; MFCUS, musculus flexor carpi ulnaris scars; PF, processus flexorius; PSD, tuberculum supracondylare dorsale;PSI,pronator superficialis insertion; SHT, sulcus humerotricipitalis; SST, sulcus scapulotricipitalis; TSV, tuberculum supracondylare ventrale. Scale bar 10 mm.

Figure 1. (A) Map of South Australia with general location of Frome Sub-Basin fossil sites marked with a rectangle, (B) detailed map of the study sites in the Frome Sub- Basin. Lake Pinpa Sites 11 and 12, Ericmas Quarry, and Wells Bog Site located by black squares.
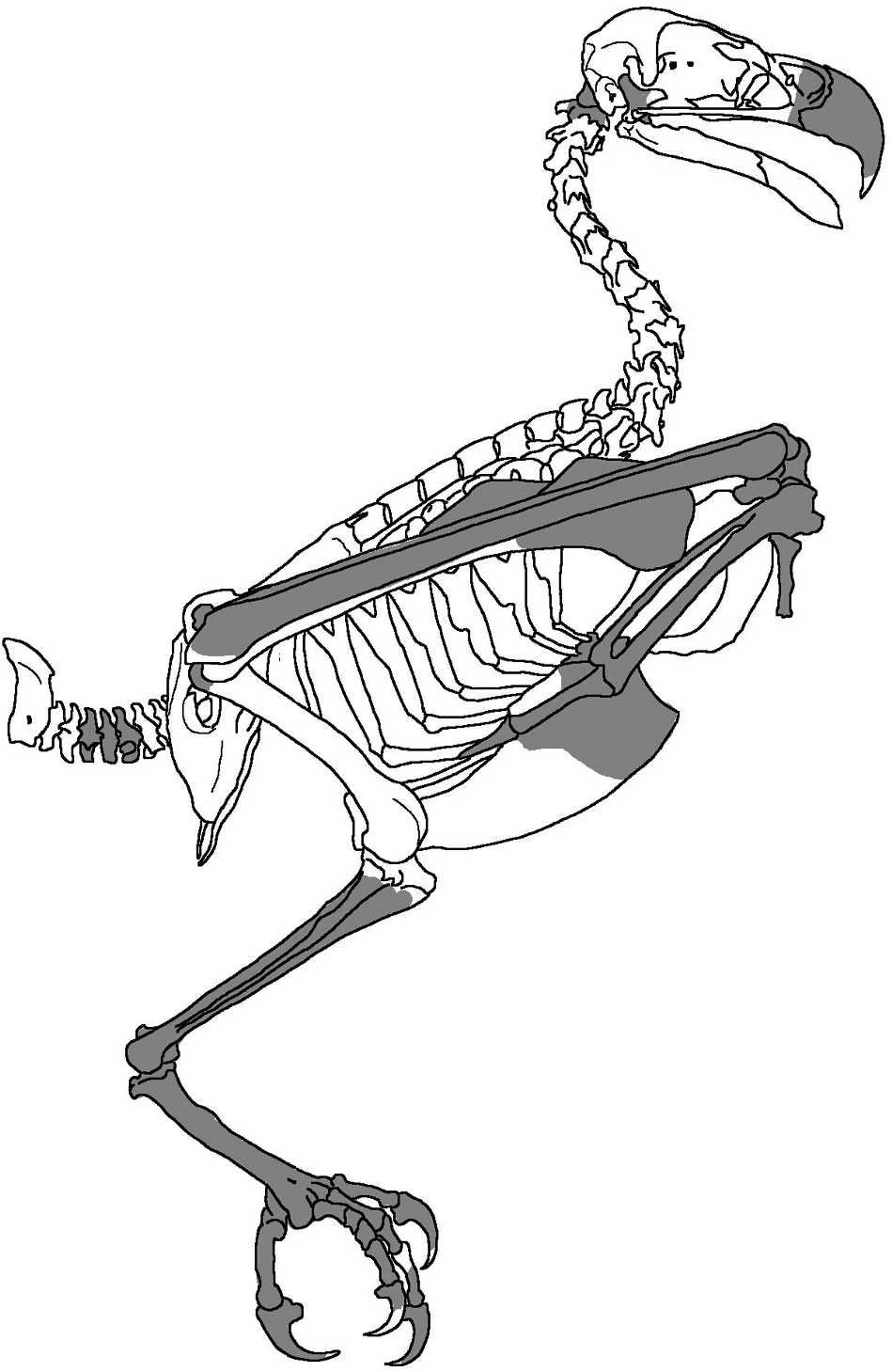
Figure 2. Exemplar accipitriform skeleton, Pandion haliaetus cristatus, derived from an illustration in Eyton (1867), showing the bones preserved in the fossil accipitrid specimen SAMA P.54998 shaded in grey. The illustrated taxon and fossil material are not identical in terms of the morphology of individual elements.

Figure 3. Archaehierax sylvestris gen. et. sp. nov. SAMA P.54998 rostrum in lateral (A) and ventral (B) view; quadrate in medial (C) and lateral (D) view; atlas vertebra in caudal (E), lateral (F) and cranial (G) view; and axis vertebra in caudal (H), lateral (I) and cranial (J) view. Specimens in A-D are coated in ammonium chloride.Abbreviations: Arc. At., arcus atlantis; CC, condylus caudalis; CM, condylus medialis; CP, condylus pterygoideus; CS, capitulum squamosum; CT, crista tomialis; FA, facies articularis; FAA, facies articularis axialis; FC, fossa condyloidea; FPB, fossa pneumaticum basiorbitale; FPC, fossa/depressio pneumaticum caudomediale; IAC, incisura caudalis arcus; IF, incisura fossae; IV, incisura ventromedialis; N, nasale; PAC, processus articularis caudalis; PM, pars maxillaris palatini; POr, processus orbitalis; POt, processus oticus. Scale bars are 10 mm.

Figure 4. Archaehierax sylvestris gen. et. sp. nov. SAMA P.54998 partial sternum in cranial (A), left lateral (B) and dorsal (C) view; right scapula in lateral (D) and medial (E) view;omal fragments of the left coracoid in medial (F) and lateral (H) view and of the right coracoid in dorsal (G) and medial (I) view. Specimens in A, B,and F-I are coated in ammonium chloride. Abbreviations: AC, apex carina; Ac, acromion; CMC, crista medialis carinae; CtS, cotyla scapularis; ColS, collum scapulae; FAC, facies articularis clavicularis; FAH, facies articularis humeralis; FoNS, foramen nervi supracoracoidei; ILA, impressio ligamenti acrocoracohumeralis; LI, labrum internum; MD, margo dorsalis; PC, pila carinae; PM, pila medialis; P. Procor., processus procoracoideus; SAC, sulcus articularis coracoideus; SE, spina externa; SMS, sulcus m. supracoracoidei; TC, tuberculum coracoideum. Scale bars are 10 mm.

Figure 12. Distal left femur NMV P.222435 depicted in cranial (A), caudal (B), medial (C) and lateral (D) view. Abbreviations: CM, condylus medialis;EL,epicondylus lateralis; FPop,fossa poplitea; IG,impressio m. gastrocnemialis lateralis;ILCC,impressio lig. cruciati cranialis; LIC, linea intermuscularis caudalis; PPAP, planum popliteum attachment point; SP, sulcus patellaris; TF, trochlea fibularis; TMGM, tuberculum muscularis gastrocnemialis medialis. Scale bar 10 mm.
IMAGES
Gallery(8)
Occurrences with images
CITATIONS
References(11)
AviList Core Team (2025) AviList: The Global Avian Checklist, v2025
Banks, R. C., R. W. McDiarmid, and A. L. Gardner, 1987: Checklist of Vertebrates of the United States, the U.S. Territories, and Canada. Resource Publication, no. 166. 79.
Benton, M.J. (ed). (1993). The Fossil Record 2. Chapman & Hall, London, 845 pp.
Benton, M.J. (ed). (1993). The Fossil Record 2. Chapman & Hall, London, 845 pp.
Bock (1994) History and nomenclature of avian family-group names