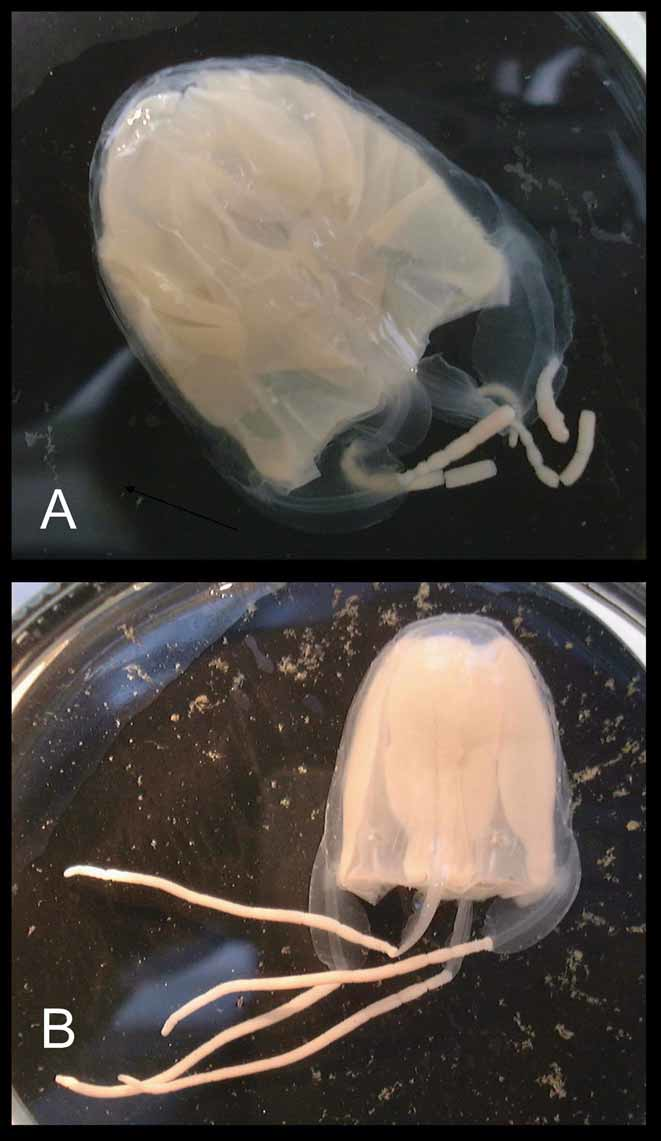
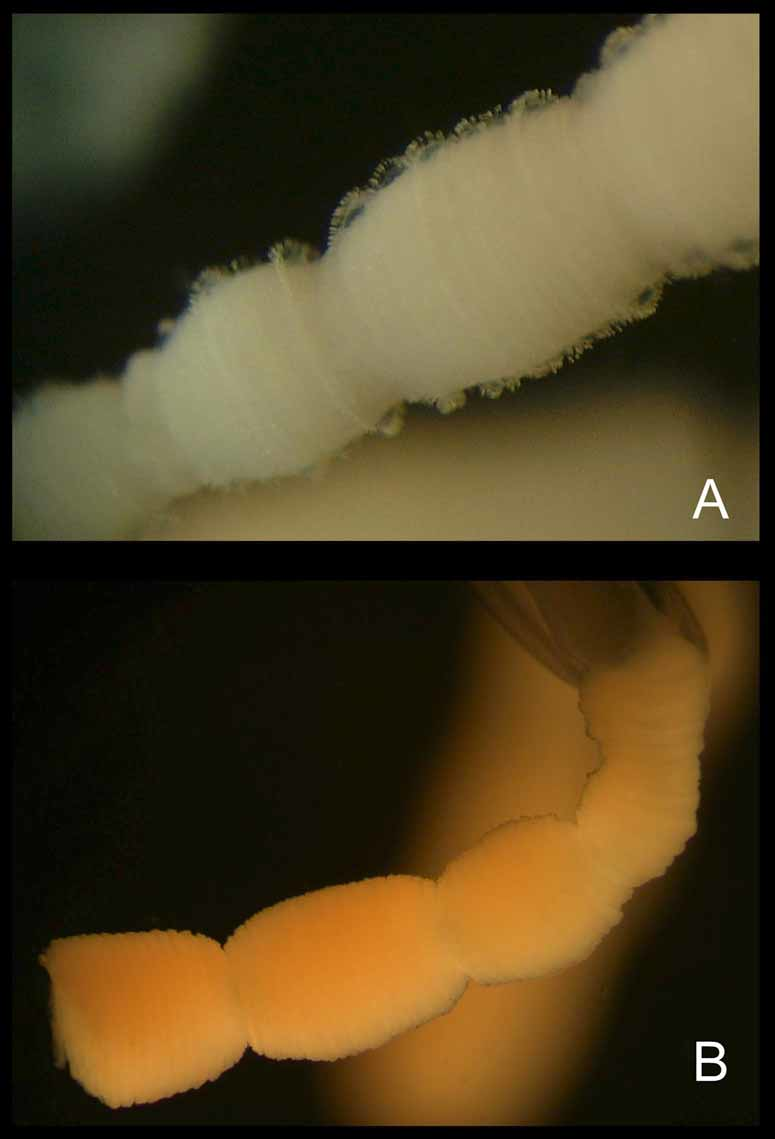
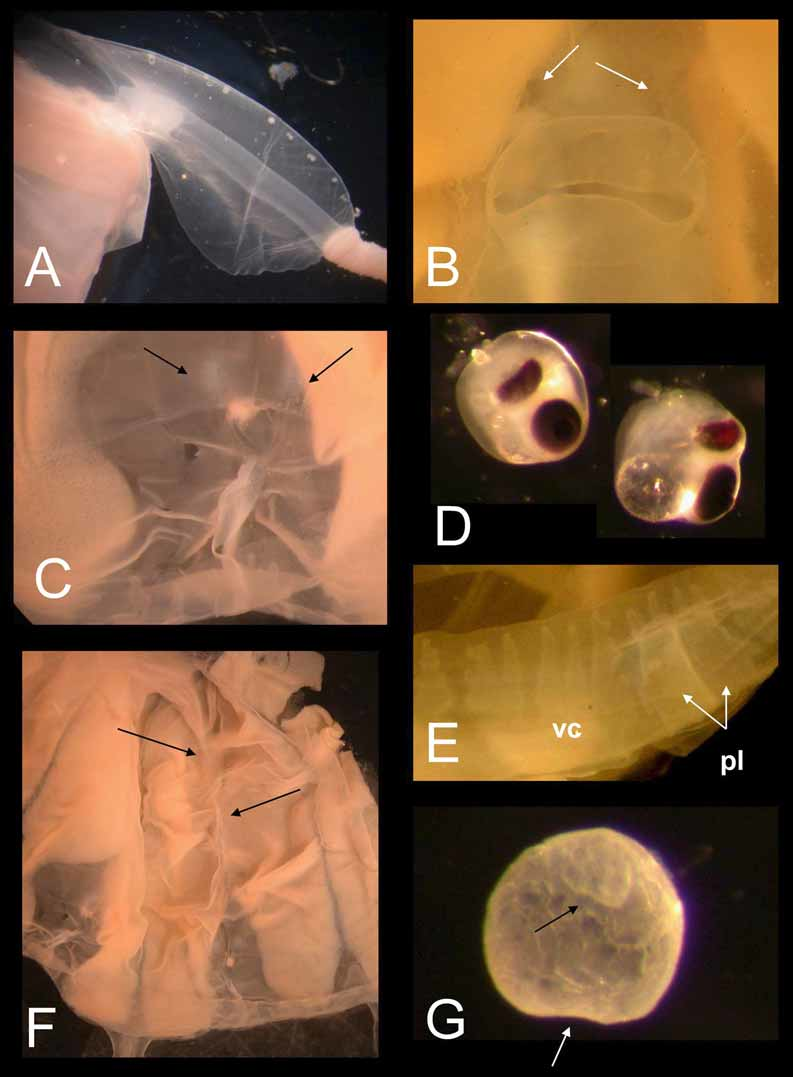
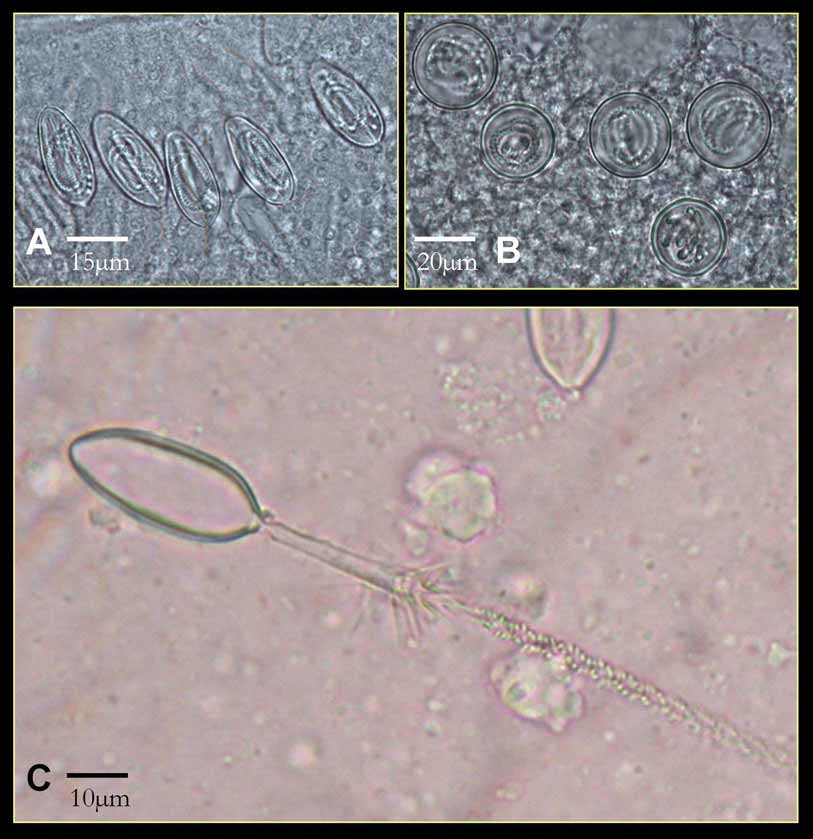
Description of holotype. Bell to about 3 cm tall, half as wide, narrower at apex than base, evenly rounded aborally (Figure 1). Mesoglea fairly thick, with the medusa typically holding its shape in water. Exumbrellar mammillations on apex; with low, round, gelatinous warts along body walls. Interradial furrows shallow, with raised ridges along either side, running the full height of bell. Adradial furrows poorly defined in upper half, shallowly defining rhopaliar region in lower half, but not defining interradial pillars. Tentacles 4, one per pedalium, appearing “ segmented ” at similar intervals, by which tentacle diameter grows distally; with halo-like thin sheet-rings perpendicular to tentacle axis, regularly spaced throughout length, approximately 60 per cm retracted (preserved); with nematocysts on halos only, inserted end on around outer edges of halos (Figure 2). Pedalia 4, interradial, long, scalpel-shaped, with moderately flared inner keel (Figure 3 A); without nematocyst freckles on outer keel. Pedalial canal about 1.40 mm wide; outer keel as wide as pedalial canal, inner keel about 3 times as wide as pedalial canal. Pedalial canals narrow and straight, with small hump near base, lacking smoothly rounded knee-like bend or upward-pointing thorn; strongly quadrate in cross section through most of length, with ridges shallower distally, unflared at tentacle insertion. Rhopalial niche ostium frown-shaped, with one upper and one lower covering scale (Figure 3 B); upper scale broadly convex with a shallow W-shape, i. e., two shallow points at about the quarter and three-quarter position. Rhopalial horns relatively short, broad, curved inward like Viking horns (Figure 3 B, C). Rhopalial windows flat, with horizontal concavity at rhopalial stalk (Figure 3 C). Rhopalial warts not observed. Eye morphology and statolith shape not discernable in preserved holotype specimen; lateral eye spots lacking in other specimens (Figure 3 D). Velarial canals palmate, with about 4 – 5 main somewhat-branched fingers per octant (Figure 3 E), all arising from a single common root extending from gastric pouch, past velarial turnover, onto velarium; lacking nematocyst patches or warts. Perradial lappets narrow triangular (Figure 3 F), with multiple finger-like extensions reaching velarial margin; with round nematocyst patches in two rows of 3 – 4. Frenulum a single sheet, lacking gelatinous buttresses; nearly reaching velarial margin. Stomach small, flat, with short manubrium; lips broadly pointed. Phacellae lacking. Mesenteries well developed, flap-like throughout stomach region to about halfway toward rhopalium; sessile cord-like another quarter way toward rhopalium, with a transparent strip of tissue reaching rhopaliar niche region. Interradial septa simple, lacking perforations. Gonads large and leaf-like along full length of interradial septa, reaching up to stomach and down past pedalia into velarial canals, overlapping perradially but with a keyhole-shaped outline around rhopaliar niches. Variation. The specimens listed separately in the paratype section above, which lack the halo-like bands, include those colloquially called “ Pseudo-Irukandji ” by Barnes (Kinsey 1988) and Gershwin (2005 a). I believe that these specimens lacking halos are an immature form of M. kingi, rather than an entirely different species (see remarks below). In paratypes with tentacles relaxed, the halo bands are about 35 per cm (preserved, e. g., paratypes SAM H 968, AM G 16005, MTQ G 55276). Curiously, the holotype and some of the paratypes have low gelatinous warts on the body, while the other paratypes lack any trace of such warts. Two pedalia on the holotype are branched, in a fashion similar to the uppermost, unpaired tentacles of the chirodropids; I believe this is just an aberration (it is lacking in other specimens of this species, but I have seen it on rare occasions in other taxa). Also in the holotype, the halo-like rings do not extend the full length of the tentacles, but are most clearly exhibited proximally. Despite these apparent aberrations, I believe this is the best specimen to represent the species because of its spectacular preservation of the peculiar endwise insertion of nematocysts, which have been somewhat worn away in the paratypes. Many of the specimens bear tentacles that are curiously “ segmented ” in appearance, much like a string of sausages; whether this is an artifact is still unclear. Some specimens have small, round nematocyst “ freckles ” on the outer keel of the pedalia, whereas the holotype does not. Most of the halo-form paratypes and some of the non-halo-form paratypes have complexly branched velarial canals, i. e., with lateral diverticula, whereas in others the canal branches are more digitiform. The taxonomic significance of these differences, if any, is not at this time well understood. Statoliths. Several statoliths have been examined from paratypes and non-type specimens of Malo kingi, and are different from those of other cubozoans in general morphology. In M. kingi, the statolith is globular in shape, with a “ basal ” indentation and an “ apical ” tooth (Figure 3 G). In the genus Carukia, for which Malo has previously been mistaken, the statoliths are sub-spherical, lacking both the tooth and “ basal ” indentation. A comparison of statolith shapes in cubozoans was given by Gershwin (2005 a). Colouration. In life, the bell is transparent and colourless; tentacles faintly pink; exumbrellar nematocyst warts pale purple. Nematocysts. As previously discussed (Gershwin and Alderslade 2005; Gershwin 2005 a; Gershwin 2005 b; Gershwin 2006 a), the principal tentacular nematocysts of Malo kingi are elongate club-shaped Type 4 microbasic p - mastigophores with a cylindrical shaft and terminal distally-aimed spines (Figure 4 A, C). This type of nematocyst is characteristic of the “ Pseudo-Irukandji ” clade of jellyfishes (Gershwin 2005 a). I found no difference in nematocyst shape between the halo-form and non-halo-form specimens, but nematocyst size differed slightly, possibly due to maturity (Halo: 32.98 – 37.56 µm long by 11.65 – 16.36 µm wide, n = 30, Non-halo: 30.27 – 36.68 µm long by 13.02 – 16.04 µm wide, n = 44). The exumbrellar nematocysts are entirely spherical isorhizas, 20.10 – 24.87 µm diameter, n = 45 (Figure 4 B). Spherical isorhizas are similar throughout the carybdeid species with exumbrellar nematocysts.
Malo kingi: A new species of Irukandji jellyfish (Cnidaria: Cubozoa: Carybdeida), possibly lethal to humans, from Queensland, Australia