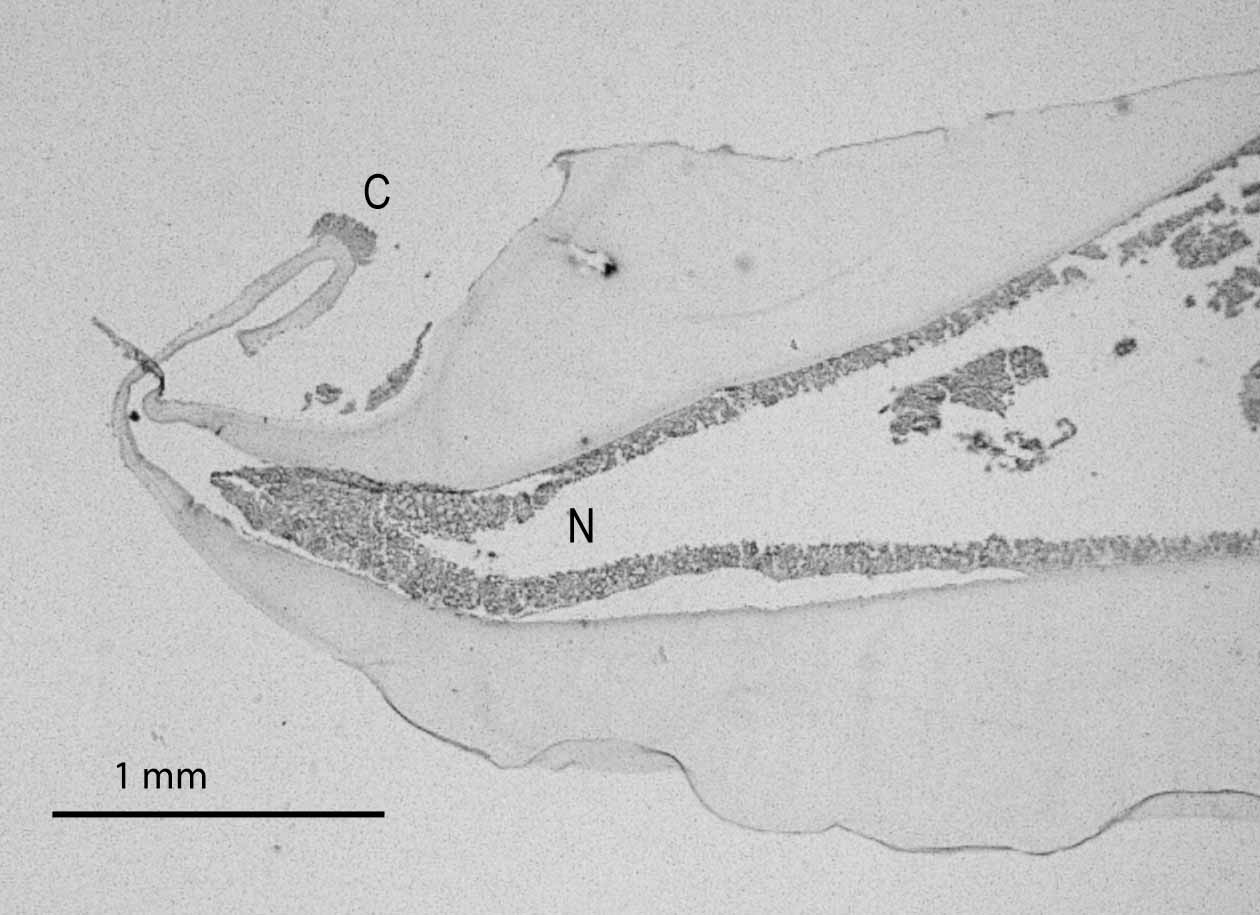
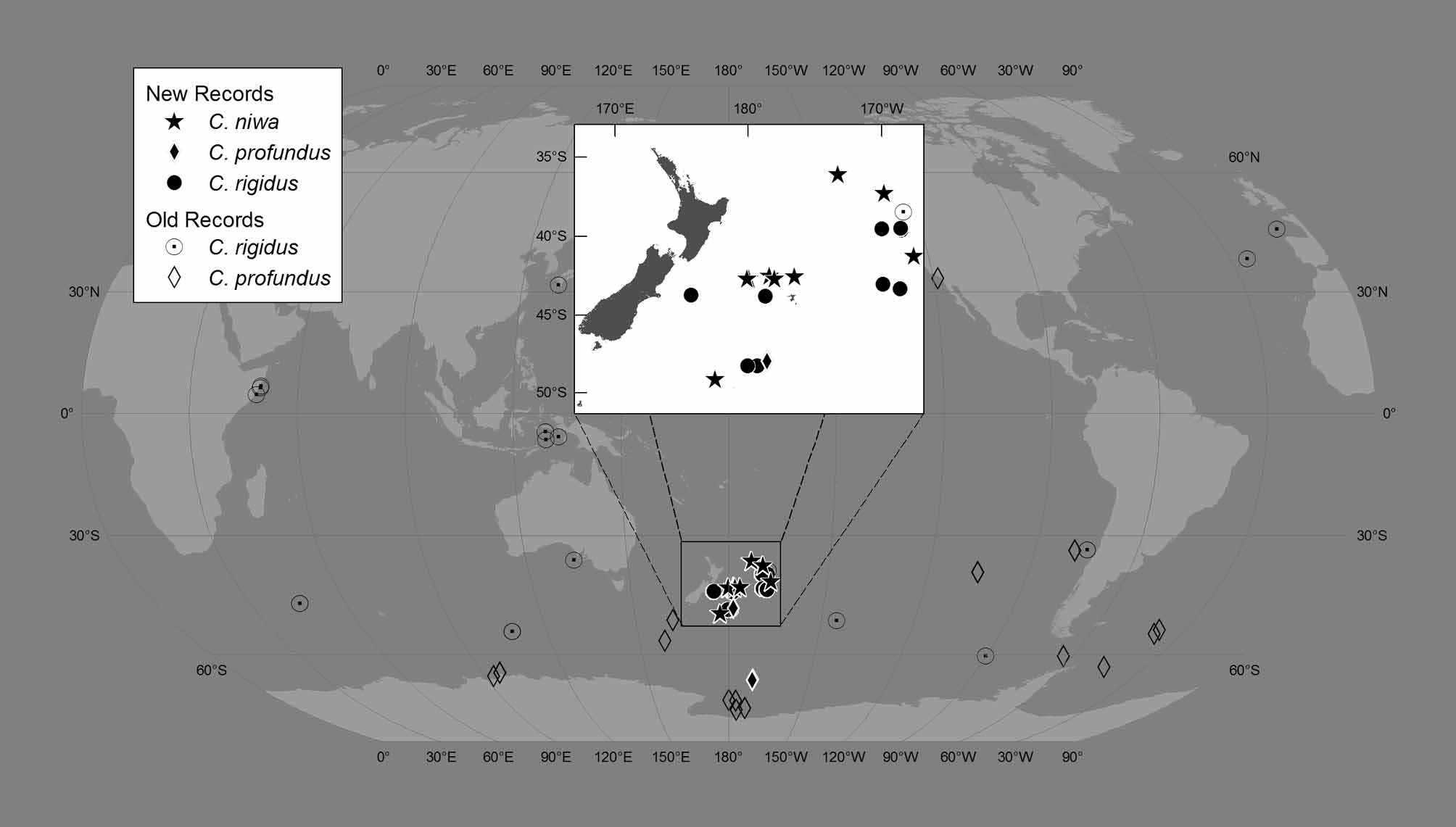
Material examined. See Table 1. Form, size, and color. Oral disc more or less circular, flat to strongly domed (Fig. 1 A), diameter 30 to 125 mm in those studied; wider than or about the same diameter as pedal disc (Fig. 1 B), which is more or less circular and flat or slightly concave. Column very short (10 – 30 mm), either straight or curved inward, so mid-column is narrower than either oral or pedal disc; typically margin overhangs pedal disc. Limbus may form a ridge as if mesoglea were bunched up by shortening of column or pulling in of pedal disc. Texture of stiff gelatin due to extraordinarily thick, hyaline mesoglea of column and oral disc (Fig. 2); pedal disc so thin that mesenterial insertions can be seen through it (Fig. 1 B). Animal 125 mm diameter (NIWA 34507: the holotype) with column and oral disc mesoglea 5 mm thick, pedal disc mesoglea 2 mm thick. Immediately after animal is collected, oral disc ectoderm brownish, rust red along mesenterial insertions; longitudinal red lines (presumably along mesenterial insertions) may be present on column. Pigment may be retained in preservation (Fig. 1 A), ectoderm may be yellowish (Fig. 1 C), or ectoderm may be entirely sloughed off. Oral disc and tentacles. Mouth central, slit-like (in holotype, mouth 18 mm long). Distal end of actinopharynx exposed, red in animals that retain pigmentation. Tentacles not contractile, short, most with acrosphere slightly greater in diameter than distal end of tentacle stalk (Fig. 1 A) (acrosphere may have detached from tentacles seeming to lack one; fragility of the stalk-acrosphere junction was noted by Fautin et al. 2002). Discal and marginal tentacles about equal in number (holotype has 56 discal, 52 marginal; paratype KUIZ 0 0 321 has 24 discal, 24 marginal; paratype NIWA 14329 has 38 discal, 38 marginal). FIGURE 1. Specimens of Corallimorphus; all except D are preserved. A, Oral disc of holotype of Corallimorphus niwa n. sp. (NIWA 34507), diameter 125 mm. B, Pedal disc of holotype of Corallimorphus niwa n. sp., diameter 100 mm. C, Paratype of Corallimorphus niwa n. sp. (ex NIWA 41750), oral disc 90 x 100 mm. D, Corallimorphus niwa n. sp. in situ. Diameter about 200 mm; location 42.64458333 º S, 177.8605833 º E; depth 1210 m. E, Corallimorphus profundus Moseley, 1877 (NIWA 41760). F, Corallimorphus rigidus Moseley, 1877 (NIWA 34571); diameter 50 mm. Discal tentacles radially and cyclically arrayed, to about 5 mm long. Most tentacles arise through very broad bump as much as 12 mm diameter, with core of endoderm visible through mesoglea (Fig. 2), giving tentacle the appearance of a nipple. Bumps more prominent in larger individuals, and within an individual along lower-order endocoels (in one individual, bumps of tentacles communicating with primary endocoels 8 – 10 mm diameter, with secondaries 8 mm, with tertiaries 5 mm). Three or four tentacles connect with each primary endocoel in largest individuals examined (e. g. holotype), highest-order endocoel with only one very small discal tentacle. Bumps diminish in size radially from mouth toward margin, and most peripheral discal tentacles lack bump. Twelve thickenings immediately around mouth ovoid, radially arrayed, lack tentacles (Fig. 1 A). Reddish radial lines on oral disc follow mesenterial insertions: 6 pairs extend from each side of mouth (along complete mesenteries, presumably), other lines extend from margin only partway to mouth; surround bumps that form base of tentacles (Fig. 1 A). Marginal tentacles very short (1 – 2 mm, each with acrosphere 1 – 1.5 mm diameter in largest individuals examined; Fig, 1 A, B, Fig. 2), may lack basal bump. Alternate ones communicate with exocoels and endocoels, the latter more orad than former. Internal anatomy. Interior poorly preserved in all specimens examined; perhaps thick mesoglea prevents penetration of preservative. Mucusy when cut. Actinopharynx with many deep longitudinal folds; ectoderm, if present, usually reddish-brown. No siphonoglyphs. Because endoderm disintegrated, nature of musculature and pattern of gametogenic mesenteries could not be determined. About 48 pairs of mesenteries, seemingly regularly arrayed. Arrangement and completeness of mesenteries could not be determined. No marginal sphincter muscle (Fig. 3). Cnidae. Cnidom spirocysts, microbasic b - mastigophores, hoplotelic microbasic p - mastigophores, holotrichs (Fig. 4). For distribution and sizes, see Table 2. This inventory may be incomplete due to poor preservation of the specimens examined. Because ectoderm is very sparse or absent on oral disc and tentacles, measurements are given for the two together. It appears, from the few samples that could be obtained from acrospheres, most of which lack ectoderm or were even entirely missing, that nematocysts of some types are confined to acrospheres. Nematocysts examined in smears of the mucusy material in the coelenteron are assumed to be from mesenterial filaments, but that could not be ascertained; the pear-shaped cnidae I refer to as holotrichs, and that are the only type of nematocyst in the actinopharynx, appeared in some smears of mesenterial filaments. Habitat. Three lines of evidence indicate the animals probably live unattached on soft sediment. An in-situ photograph (Fig. 1 D) of what is almost certainly Corallimorphus niwa n. sp., based on length and distribution of capitate tentacles, was taken in an area of muddy soft sediment. The pedal discs of all individuals examined were unattached, and showed no evidence of having been attached to anything; specimens of some other species of Corallimorphus (Fautin 1984; below) had pebbles attached to the pedal disc. The low discoidal form of the animal is hydrodynamically favorable for an unattached polyp; it is similar to that of the sea anemone Liponema, which lives unattached on soft sediments (Dunn & Bakus 1977). Type locality. Northern edge of Chatham Rise (42.5390 – 42.5433 º S, 178.3382 – 178.3400 º E), at 1404 – 1414 m. Geographic and bathymetric distribution. Corallimorphus niwa n. sp. is known only from immediately around New Zealand (Fig. 5), at depths of about 900 to 1700 m.
Corallimorphus niwa new species (Cnidaria: Anthozoa), New Zealand members of Corallimorphus, and redefinition of Corallimorphidae and its members