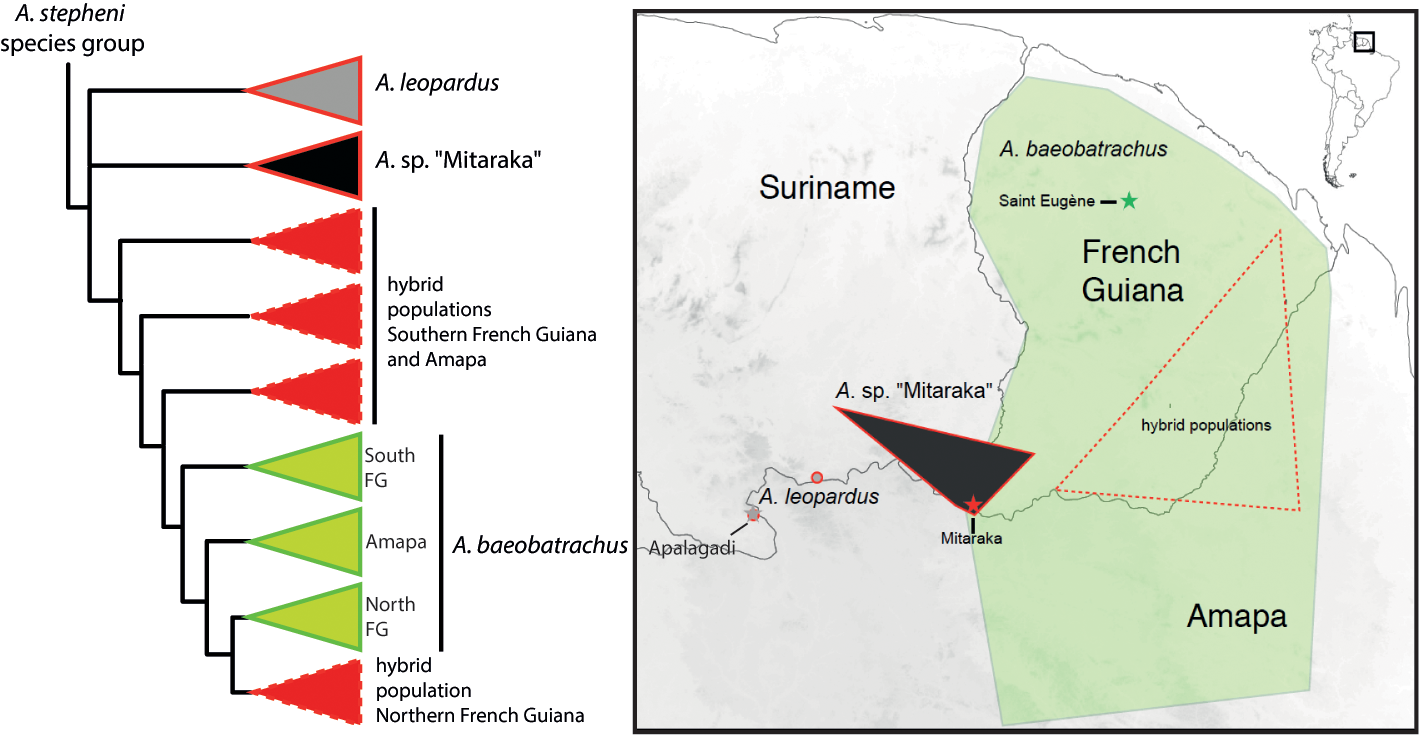
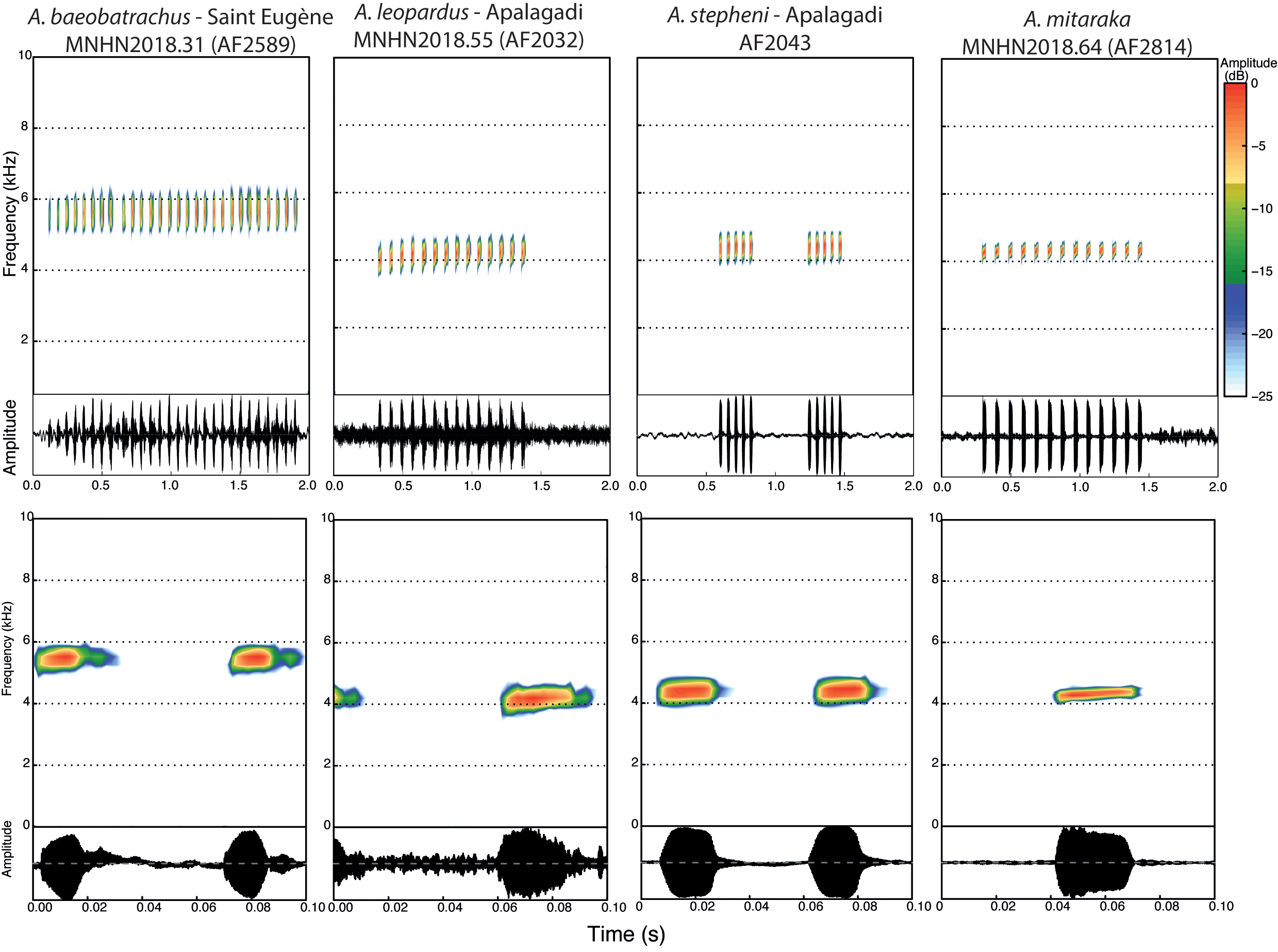
Morphological comparisons with other Anomaloglossus. The only other species group co-occurring with the Anomaloglossus stepheni group is the A. degranvillei group, currently represented by A. blanci Fouquet, Vacher, Courtois, Villette, Reizine, Gaucher, Jairam, Ouboter & Kok, 2018, A. degranvillei (Lescure, 1975), A. dewynteri Fouquet, Vacher, Courtois, Villette, Reizine, Gaucher, Jairam, Ouboter & Kok, 2018, and A. surinamensis Ouboter & Jairam, 2012, which are readily distinguishable by their moderate webbing (basal in A. baeobatrachus) and well-developed fringes on all toes (smaller and more developed on Toe II, III, IV) and the absence of a dorsolateral stripe (present). Outside these two groups all the other described Anomaloglossus species exclusively occur in Pantepui (i. e., the Guiana Shield highlands, see Kok et al. 2018) and have moderate to extensive toe webbing except A. meansi Kok, Nicolaï, Lathrop & MacCulloch, 2018, A. kaiei (Kok, Sambhu, Roopsind, Lenglet & Bourne, 2006), A. rufulus (Gorzula, 1990) and A. roraima (La Marca, 1997). However, unlike A. baeobatrachus, these last four species lack developed fringes on toes. Within the Anomaloglossus stepheni group, A. baeobatrachus can be distinguished from A. stepheni (Figs. 2 – 3, Table 1) by (1) skin of dorsum irregularly tuberculate in A. baeobatrachus vs. evenly tuberculate in A. stepheni; (2) ventral coloration with yellow shades, at least posteriorly in males in A. baeobatrachus vs. entirely translucent white in A. stepheni; (3) dorsolateral line thin and interrupted in A. baeobatrachus vs. thick and ill-defined margins dorsally in A. stepheni; (4) call much longer (mean = 1.12, range 0.84 – 1.33 s) in A. baeobatrachus [n = 9] vs. mean = 0.25, range 0.18 – 0.29 s in A. stepheni [n = 4]) emitted at a slower pace (call rate mean = 0.33, range 0.18 – 0.48 calls / s in A. baeobatrachus [n = 11] vs. mean = 1.40, range 1.10 – 1.89 calls / s in A. stepheni [n = 4]), with a higher dominant frequency (mean = 5.39, range 4.96 – 5.59 kHz in A. baeobatrachus [n = 6] vs. mean = 4.48, range 4.30 – 4.85 kHz in A. stepheni [n = 4]). Anomaloglossus baeobatrachus can be distinguished from A. leopardus (Figs. 2 – 3, Table 1) by (1) a smaller body size (mean = 16.2; range 14.8 – 17.1 mm in males [n = 16] and mean = 18.4; range 17.3 – 19.4 mm in females [n = 4] in A. baeobatrachus vs. mean = 18.8, range 18.5 – 19.2 mm in males [n = 10] and mean = 21.1, range 20.5 – 21.5 mm in females [n = 4] in A. leopardus); (2) dorsal coloration uniformly dark brown with inconspicuous dark transverse bars on legs in A. baeobatrachus vs dorsal coloration with large dark blotches, particularly visible in preservative and conspicuous dark transverse bars on legs in A. leopardus; (3) ventral coloration with yellow restricted to the posterior part in males in A. baeobatrachus vs. generally entirely yellow in A. leopardus; (4) dorsolateral line thin and discontinuous in A. baeobatrachus vs. thick and continuous in A. leopardus; (5) call with a faster note rate (mean = 16.1, range 15.7 – 16.8 note / s in A. baeobatrachus [n = 9] vs. mean = 13.4, range 12.9 – 13.6 notes / s in A. leopardus [n = 4]) with a higher dominant frequency (mean = 5.39, range 4.96 – 5.59 kHz in A. baeobatrachus [n = 6] vs. mean = 4.49, range 4.40 – 4.57 kHz in A. leopardus [n = 4], Table 2). Description of the neotype. An adult male, 15.9 mm SVL; body robust; head wider than long, HL 94 % of HW; HL 32 % of SVL; dorsal skin irregularly tuberculate, one enlarged tubercle on each eyelid, snout long (SL 55 % of HL), rounded to nearly truncate in dorsal view, protruding in lateral view, extending past lower jaw. Nares located anterolaterally; canthus rostralis rounded, loreal region concave; IN 44 % of HW; EN 31 % of HL, 70 % of ED. Tympanum distinct anteroventrally; supratympanic fold diffuse; choanae small, circular, located anterolaterally to palatine (Fig. 3). Forelimb slender, skin dorsally tuberculate; metacarpal ridge present; HAND 24 % of SVL; Finger I longer than Finger II when fingers adpressed; fingers large and flattened without webbing, lateral fringes present on preaxial edges of Fingers II; Finger III distinctly swollen dorsally and preaxially; tip of Finger IV not reaching distal subarticular tubercle on Finger III when fingers adpressed; finger discs expanded, wider than long, about 1.5 X width of digit; width of disc on Finger III 0.5 mm; discs with distinct dorsal scutes. Relative lengths of adpressed fingers III> IV> I> II; palmar tubercle large, heart-shaped, 0.7 mm in diameter (larger than Finger III disc), thenar tubercle small (equal to Finger III disc), elliptic, half the size of palmar tubercle, well separated from palmar tubercle. Only basal subarticular tubercles on Fingers I, II and IV are conspicuous; Subarticular tubercle of Finger I larger than Finger II and IV subarticular tubercles. Hind limb robust, skin dorsally tuberculate; TL 47 % of SVL; heels not in contact when hind limbs are flexed at right angle to sagittal plane of body; FL 42 % of SVL; relative length of adpressed toes IV> III> V> II> I; Toe I very short, its tip reaching the base of subarticular tubercle on Toe II when toes adpressed; toe discs larger than width of toes; disc on Toe I only slightly larger than width of digit. Width of disc on Toe IV 0.7 mm. Feet poorly webbed; lateral fringes present on all toes. Toe webbing formula I 1 + – 1 - II 1 + – 1 - III 1 ½ – 1 ½ IV 0 – 1 + V. One to three subarticular tubercles on toes as follows: one on Toes I and II, two on Toes III and V, three on Toe IV. Inner metatarsal tubercle protuberant elliptical, 0.5 mm in length, outer metatarsal tubercle round, protuberant, 0.3 mm in diameter. Tarsal keel well defined, tubercle-like and strongly curved at proximal end. Metatarsal fold conspicuous and strongly curved. Color of neotype in life. Dorsal color uniformly dark brown. Discontinuous pale dorsolateral stripes consisting of irregular bluish, white, orangish and yellow (near groin) small blotches (Fig. 2). Dark brown lateral band extending from tip of snout to the groin and containing the indistinct dorsal part of tympanum. Upper lip with small iridescent light blue blotches. Lower flanks pale yellow with small iridescent white blotches. Throat anteriorly yellow, centrally white and posteriorly pinkish, covered with melanophores more densely anterolaterally; belly centrally white and laterally and posteriorly yellow, ventral surface of thighs and arms pale orange. Iris with copper metallic pigmentation and pupil ring interrupted dorsally and ventrally by transversal dark pigmentation (Fig. 2). Upper arm pale orange dorsally with a conspicuous proximal white spot, anteriorly and posteriorly light brown with small bluish spots. Lower arm light brown with ill-defined dark brown blotches and small bluish spots. Dorsal surfaces of thigh, shank and tarsus dark brown with ill-defined darker blotches. Paracloacal marks orangish, elongated anteroposteriorly. Toes and digits with small light blue dots. Palms and soles dark brown. Color of neotype in preservative. After three years in 70 % ethanol, some colors of the specimen faded and the dorsal coloration now varies from brown to grey and the yellow ventral coloration disappeared. All bluish freckles and orange and reddish marks turned cream (Fig. 3). Variation. Measurements (range, mean, and standard deviation) are provided in Table 1. Adult dorsal coloration varies from brown to reddish brown in males, females being paler and with more conspicuous darker dorsal blotches. The dorsolateral line varies from almost continuous to continuous; bluish flecks may be numerous or absent. Additionally, overall coloration and tuberculation may vary with light intensity, time of the day and probably reproductive activity as males carrying tadpoles apparently display overall lighter colors, smoother skin and sharper contrasts while calling males are very dark and highly tuberculate. Ventral coloration of female is entirely yellow while the yellow parts are limited in various extent to the posterior region and throat in males. Vocal sac, slits and small dark melanophores on throat only observed in males as well as swelling on the Finger III. Advertisement call. Nine specimens (two not collected) calling from the leaf litter were recorded from a distance of about 1 m and at temperatures ranging from 23 to 26 ° C. Descriptive statistics of call parameters are presented in Table 2. Anomaloglossus baeobatrachus emits trains (call length mean = 1.12 s; range 0.84 – 1.33 s) of short notes (note length mean = 0.019 s; range 0.018 – 0.022 s; inter-note interval mean = 0.045 s; range 0.043 – 0.046 s) (Fig. 4). The spectral structure of the note has a developed harmonic structure and the dominant frequency is 5.39 kHz on average (range 4.96 – 5.59 kHz) with a slight upward modulation (ca. 0.2 kHz) (Fig. 4, Table 2). Larval morphology. The following description is based on four tadpoles at stage 30, 32, 33 and 34 (Fig. 5). Measurements are presented in Table 3. Tadpoles correspond to Type 2 tadpoles of Orton (1953) although an external spiracle is absent; tadpoles endotrophic; body skin smooth; TL 10.86 – 12.92 mm; BL 3.97 – 4.62 mm, 36 – 37 % of TL, 141 – 159 % of BW, 212 – 241 % of BH; BW 137 – 171 % of BH (Table 2); body inflated from trophic reserves until stage 33 when it becomes ovoid; round snout in dorsal and lateral view; eyes positioned and directed laterally (Fig. 5); ED 0.46 – 0.68 mm, 65 – 81 % of IOD; IOD more than IND; nares positioned frontally and directed laterodorsally; narial opening reniform in lateral view; END 0.2 8 – 0.40 mm. Spiracular tube absent. Lateral-line system inconspicuous. Caudal musculature highest at its base, tapering posteriorly, terminating at tail tip; tail tip rounded; upper fin originating at junction of body and tail, gradually increasing in height to about 3 / 4 of tail; UTF 39 – 50 % of TMH; LTF 26 – 42 % of TMH; MTH 15 – 18 % of TL. Mouth small and bare (without marginal papillae, labial teeth nor sheaths), directed ventrally, oral disc width 0.5 mm (from edges of jaws) (Fig. 5, Table 3). In life, until stage 32 the entire body is light grey except the tail fins that are translucent and the abdomen that is yellowish. From stage 33 the body becomes dark grey with golden speckles, and the tail and fin become translucent posteriorly (Fig. 5). Tadpoles of Anomaloglossus baeobatrachus can be distinguished from those of any other Anomaloglossus of the stepheni group (except A. apiau and A. stepheni) by the absence of functional mouth and the prominent trophic reserves. Only two other Anomaloglossus with an endotrophic tadpole have been described, A. stepheni (Juncá et al. 1994) and A. degranvillei (Lescure 1984); although the latter may in fact correspond to A. surinamensis or A. blanci (Fouquet et al. 2018). Tadpoles of A. baeobatrachus can be distinguished from those of Anomaloglossus stepheni by the absence of a spiracle (present in A. stepheni), a mouth located anteriorly to the eyes (at the level of the eyes in A. stepheni), eyes located dorsolaterally (laterally in A. stepheni), a proportionally shorter tail – BL 36 – 37 % of TL (vs. 33 % in A. stepheni) (Juncá et al. 1994). They can be distinguish from tadpoles of A. degranvillei by the presence of an oral disc (absent in A. degranvillei), presence of a vent tube (absent in A. degranvillei), eyes located dorsolaterally (laterally in A. degranvillei) and by their development, which is not completed on the back of the male parent.
A new species of Anomaloglossus (Anura: Aromobatidae) of the stepheni group with the redescription of A. baeobatrachus (Boistel and de Massary, 1999), and an amended definition of A. leopardus Ouboter and Jairam, 2012