Anomaloglossus
Anomaloglosse(+2)·Grant, Frost, Caldwell, Gagliardo, Haddad, Kok, Means, Noonan, Schargel & Wheeler, 2006
GBIF:2426205
0
Descendants
0
Children
0
Species
PROFILE
Species Profile
Characteristics
ABOUT
Descriptions(11)
Eastern Pantepui Anomaloglossus species
Anomaloglossus megacephalus can be distinguished from A. beebei by (characters of A. beebei in parentheses, see also Kok et al. 2006b and Kok & Kalamandeen 2008) its larger size, female SVL max 28.3 mm [n=3] in A. megacephalus (18.7 mm in A. beebei, n=27), fingers with keel-like lateral folds, best developed preaxially on Fingers II and III (fringes not folded), toes with folded flaplike fringing (fringes not folded), throat in adult female blotched (immaculate), palm dark brown to black (yellowish), distinct dark bands on thigh and shank (absent), dark interorbital V-shaped band (absent).
Anomaloglossus megacephalus can be distinguished from A. breweri (Barrio-Amorós, 2006) by (characters of A. breweri in parentheses) its larger size, female SVL max 28.3 mm [n=3] in A. megacephalus (23.8 mm in A. breweri, n=1), throat in adult female blotched (immaculate), median lingual process short, wider than long, tapered (distinctly longer than wide, tip pointed).
Anomaloglossus megacephalus can be distinguished from A. kaiei by (characters of A. kaiei in parentheses, see also Kok & Kalamandeen 2008) its larger size, female SVL max 28.3 mm [n=3] in A. megacephalus (19.8 mm in A. kaiei, n=25), fingers with keel-like lateral folds, best developed preaxially on Fingers II and III (fringes not folded), toes moderately webbed (basally webbed), dorsolateral stripe absent (present), oblique lateral stripe present (absent), throat in adult female blotched (immaculate).
Anomaloglossus megacephalus can be distinguished from A. murisipanensis (La Marca, 1998) in having (characters of A. murisipanensis in parentheses, all based on the preserved holotype, which is a juvenile and the only known specimen) fingers with keel-like lateral folds, best developed preaxially on Fingers II and III (fringes barely detectable), more webbing on toes, symmetrical cloacal tubercles present (absent), tarsal keel weakly to distinctly curved, slightly tuberclelike (straight, not tuberclelike), oblique lateral stripe always present, even if broken in spots (not distinguishable), no white stripe between naris and eye in preservative (present).
Anomaloglossus megacephalus can be distinguished from A. praderioi by (characters of A. praderioi in parentheses, see also Kok 2010) its larger size, female SVL max 28.3 mm (n=3) in A. megacephalus (22.7 mm in A. praderioi, n=1), Finger IV longer than I (equal), tip of Finger IV always surpassing the base of the distal subarticular tubercle on Finger III when fingers adpressed (barely reaching the base), toes moderately webbed with folded flaplike fringing (basally webbed, folded flaplike fringing not present on all toes), dorsolateral stripe absent (present), oblique lateral stripe present (absent), throat in adult female blotched (immaculate).
Anomaloglossus megacephalus can be distinguished from A. roraima in having (characters of A. roraima in parentheses) fingers with keel-like lateral folds, best developed preaxially on Fingers II and III (fringes barely distinct, not distinctly folded), all toes with folded flaplike fringing (absent), webbing on toes moderate (absent).
Anomaloglossus megacephalus can be distinguished from A. rufulus (Gorzula, 1990) in having (characters of A. rufulus in parentheses) webbing on toes moderate (rudimentary), and most remarkably by ventral colour pattern; light with a few dark brown blotches in A. megacephalus (dark brown marbled with white blotches in A. rufulus).
The new species superficially most resembles A. parkerae, A. tepuyensis (with which it has been confused), and A. triunfo (Barrio-Amorós, Fuentes-Ramos & Rivas-Fuenmayor, 2004), in sharing a similar dorsal pattern. It should be noted that A. triunfo is a possible synonym of A. tepuyensis (Kok & Barrio- Amorós unpubl. data, see also comments by Myers & Donnelly 2008). Anomaloglossus parkerae is distinguished from A. tepuyensis and A. triunfo only by a few characters such as size, definition of dorsal pattern, and condition of finger and toe fringes (Myers & Donnelly 2008). Adults of A. megacephalus can notably be distinguished from those three similar taxa in having a more slender body ( Anomaloglossus parkerae, A. tepuyensis and A. triunfo are distinctly stockier frogs), a comparatively longer snout and a larger, somewhat more massive head [See Fig. 5 for close comparison between A. megacephalus and A. tepuyensis and A. parkerae; compare also with Duellman’s (1997: 10) colour plate of A. parkerae, fig. 30 (showing A. parkerae) in Lötters et al. (2007), and figs. 13, 15 and 16 illustrating A. tepuyensis in Myers & Donnelly (2008); see also below for further comments], and in having symmetrical cloacal tubercles (absent in all females of A. parkerae, A. tepuyensis and A. triunfo examined).
Direct comparison of specimens shows a difference in head size in Anomaloglossus megacephalus compared to the similar A. parkerae, A. tepuyensis and A. triunfo, as confirmed by plotting of morphometric proportions. Comparison of HL3 with BEL indicates that the new species proportionally has a longer head than A. tepuyensis (Fig. 6). In order to distinguish between those similar species, the midline distance from the tip of the snout to an imaginary line between anterior insertions of upper arms is measured, and that distance is then extended posteriorly from the same imaginary line. In two of the three available A. megacephalus adult specimens the distance reaches or surpasses the anterior insertion of thigh, whereas in all adult specimens of A. parkerae, A. tepuyensis and A. triunfo examined (n = 20), the distance fails to reach the thigh insertion (see Fig. 5 B). It is noteworthy to mention that the only specimen of A. megacephalus in which HL3 is not longer than BEL is in poor preservation state.
Export occurrence data
Darwin Core Archive (ZIP)
Common names used for this species across different languages and regions. From 1 country.
Vernacular (common) names are the everyday names used for a species in different languages and regions. A single species may have dozens of common names worldwide.
CLASSIFICATION
Taxonomic Classification Tree

HIERARCHY
Child Taxa(32)
RELATED
Related Name Usages(20)
Matching names from other GBIF-indexed checklists and datasets.
MULTIMEDIA
Media Files(9)
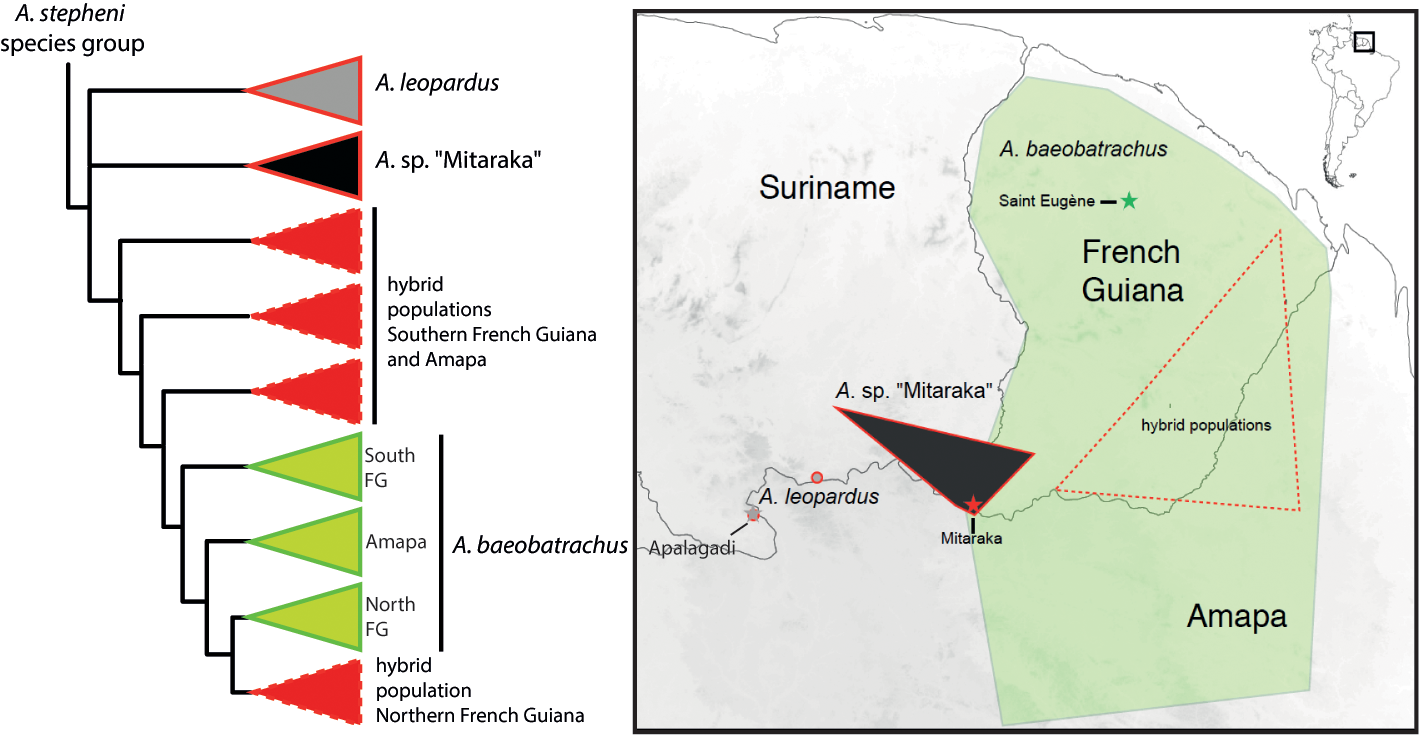
FIGURE 1. Phylogenetic tree adapted from Fouquet et al. (2019) and respective distributions of A. baeobatrachus, A. leopardus and A. sp “Mitaraka”. Red contours of collapsed branches in the tree indicate groups with exotrophic tadpoles, while green contours indicate groups with endotrophic tadpoles. Stars with corresponding colours indicate type localities.

FIGURE 2. Three males (top) and one female (bottom) of four species of the Anomaloglossus stepheni group. Size of the specimens with asterisk is not scaled.

FIGURE 3. Views of dorsum, venter, left hand, left foot and Median Lingual Process of the neotype of A. baeobatrachus (left column), of a topotypical specimen of A. leopardus (central column) and of the holotype of A. mitaraka sp. nov. (right column).
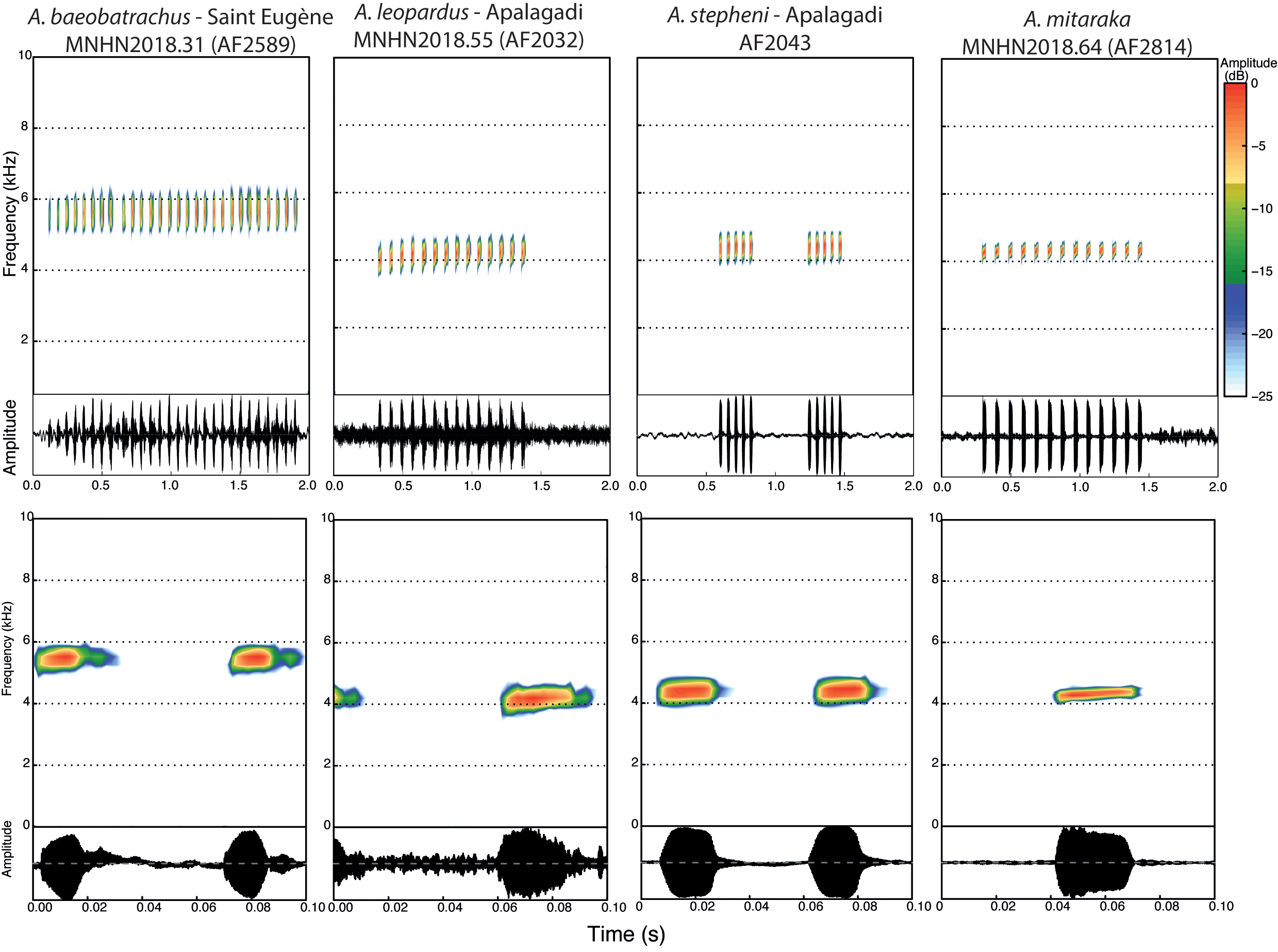
FIGURE 4. Spectrograms and oscillograms of typical call of four species of the Anomaloglossus stepheni group in a 2 s and a 0.1 s time windows.

FIGURE 6. Tadpoles of Anomaloglossus mitaraka: on the left dorsal, lateral and ventral pictures of AF2875A preserved (top) and lateral picture of AF2875B in life (bottom); on the right picture of AF2732A in the nest (top), froglet AF2875B (middle), male AF2878 carrying tadpoles to the water (bottom).

FIGURE 5. Comparison of Anomaloglossus megacephalus sp. nov. and the similar A. tepuyensis and A. parkerae. A: Dorsal view of preserved specimens of (from left to right) A. megacephalus sp. nov. (IRSNB 1986, 27.3 mm SVL), A. tepuyensis (MHNLS 17401, 23.3 mm SVL) and A. parkerae (MHNLS 11089, 23.2 mm SVL). B: Comparison between midline distance from snout tip to an imaginary line between anterior arm insertions and the same distance reported posteriorly from the same imaginary line in A. megacephalus sp. nov. and A. tepuyensis. In most A. megacephalus specimens (left, IRSNB 1986, 27.3 mm SVL) the distance reaches or surpasses the anterior insertion of thigh, whereas in all specimens of A. tepuyensis examined (right, MHNLS 17401, 23.3 mm SVL), the distance fails to reach the thigh. Photos by PJRK.
IMAGES
Gallery(9)
Occurrences with images
CITATIONS
References(6)
Article title: Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia: Athesphatanura: Dendrobatidae).
Bulletin of the American Museum of Natural History 299.
inferred from original work (title)
original publication
web search (AJR)